Федеральное агентство по образованию
Российский государственный университет
Нефти и газа имени И. М. Губкина
Кафедра органической химии и химии нефти
Курсовая работа по теме
"Свойства адамантана"
Выполнили:
Ст. гр. ХТ-08-5
Волкова В.С.
Проверил:
Ст.пр. Гируц М.В.
Москва 2010
1. Общие сведения
2. Номенклатура
3. Получение
3.1 Из природных источников
3.2 Синтетические методы
4. Физические свойства
4.1 Индивидуальное вещество
4.2 Структурные свойства
4.3 Спектральные свойства
5. Химические свойства
5.1 Адамантильные катионы
5.2 Реакции по узловым положениям
5.2.1 Бромирование
5.2.2 Алкилирование
5.2.3 Фторирование
5.2.4 Карбоксилирование
5.2.5 Гидроксилирование
5.2.6 Нитрование
5.3 Реакции по мостиковым положениям
6. Применение
7. Экспериментальная часть
Литература
адамантан углеводород синтез узловой мостиковый
1. Общие сведения
Адамантан – это химическое соединение, насыщенный трициклический мостиковыйуглеводородсформулойC 10 H 16 .Молекула адамантана состоит из трёхциклогексановыхфрагментов, находящихся в конформации "кресло". Пространственное расположение атомовуглеродав молекуле адамантана повторяет расположение атомов вкристаллической решёткеалмаза. Свое название адамантан получил от ἀδάμας ("непобедимый" - греческое название алмаза).
2. Номенклатура
Согласно правилам систематической номенклатуры, адамантан следует называть трициклодекан. ОднакоИЮПАКрекомендует использовать название "адамантан" как более предпочтительное. Молекула адамантана обладает высокой симметрией. Вследствие этого 16 атомовводородаи 10 атомовуглерода, образующие её, могут быть отнесены всего к двум типам.

Положения типа1называютсяузловыми, а положения типа2 -мостиковыми. В молекуле адамантана насчитывается четыре узловых и шесть мостиковых положений.
Обычно используют такие изображения структурной формулы молекулы адамантана:

Таким образом, узловые атомы углерода – 1,3,5,7, а мостиковые – 2,4,6,8,9,10.
В дизамещенных производных адамантана с одним мостиковым заместителем пространственная ориентация мостикового заместителя может быть аксиальной (а) или экваториальной (е) в зависимости от расположения заместителя по отношению к плоскости общего для обоих заместителей циклогексанового кольца или ее можно обозначить как цис- и транс- :

1. В случае отсутствия узловых заместителей нумерацию атомов углерода проводят с учетом предпочтительности заместителя таким образом, чтобы более предпочтительный мостиковый заместитель имел меньший номер, а сумма номеров углеродных атомов была минимальной. При обозначении алкиладамантанов меньший номер получает более простой заместитель.
2. При наличии одного узлового заместителя ему придается номер 1, нумерация других атомов углерода ядра при этом производится с соблюдением положений пункта 1.
3. При наличии нескольких узловых алкильных заместителей номер 1 получает узловой заместитель, более предпочтительный согласно правилам ИЮПАК.
4. Атомы углерода, получившие, согласно приведенным выше правилам, номера 1-9, составляют рациональный фрагмент бициклононана данного производного адамантана, при этом положения мостиковых заместителей углеродных атомов 2,4,6 и 8 определяются как экзо- или эндо- в зависимости от того, направлен ли заместитель соответственно вверх или вниз по отношению к плоскости рационального фрагмента бициклононана, у атома 10 – как цис- или транс- по отношению к атому 1, а у 9 – как син- или анти- в зависимости от того, направлен ли он вправо или влево относительно заместителя 1.
3. Получение
3.1 Из природных источников
В настоящее время единственным природным продуктом, содержащим адамантан и его гомологи, является нефть. Содержание этого углеводорода в нефти составляет всего 0,0001-0,03 % (в зависимости от месторождения), вследствие чего такой способ получения адамантана является экономически невыгодным. Помимо самого адамантана, в нефти присутствуют его многочисленные производные. Таких соединений известно более тридцати. Методы идентификации адамантана в нефтях и его выделения основаны на его необычных для углеводородов данной молекулярной массы свойствах: высокая температура плавления, летучесть, малая растворимость, а также способность образовывать устойчивые аддукты с тиокарбамидом.
Выделение адамантана из нефти, не имеющей бензиновых фракций, осуществляется путем однократной обработки тиокарбамидом дистиллатов, отогнанных из нефти с водяным паром. При охлаждении полученного тиокарбамидного экстракта до –50°С адамантан выкристаллизовывается и легко отделяется фильтрованием. Та получают около 75% адамантана, присутствующего в нефти.
Если в нефти имеются легкие фракции и содержание адамантана небольшое, то обработку дистиллата тиокарбамидом повторяют, используя небольшое его количество, и получают высокоселективные экстракты. Дальнейшее количественное выделение адамантана может быть осуществлено также методами препаративной ГЖХ. Для выделения адамантана из нефти может быть также использован метод азеотропной перегонки циклопарафинового концентрата с три(лерфторбутил)амином.
Выделение адамантана из парафиновых нефтей требует более эффективных методов его концентрирования, таких, как термодиффузия и препаративная ГЖХ. Как показали исследования, наилучшие результаты при выделении адамантана дает метод, сочетающий перегонку дистиллата (с перегретым водяным паром) с последующим выделением путем препаративной ГЖХ.
3.2 Синтетические методы
Первый успешный синтез адамантана из эфира Меервейна был осуществлёнВ. Прелогомв1941 году. Синтез включал несколько стадий, а выход адамантана не превышал одного процента.

Этот метод уже не используется для синтеза адамантана в связи с высокой трудоёмкостью и низким выходом конечного продукта. Однако он имеет некоторую ценность в плане получения различных производных адамантана, в частности 1,3-адамантандикарбоновой кислоты.
Для получения этого углеводорода в лабораторных условиях в настоящее время используют метод Шлейера. Димерциклопентадиена(который является вполне доступным соединением) подвергается каталитическомугидрированию, после чего изомеризуется в адамантан в присутствии катализатора - кислоты Льюиса. Методика, описанная в Organic Syntheses, предусматривает использованияоксида платиныв качествекатализаторагидрирования, а такжехлорида алюминияв качестве катализатора изомеризации.При этом выход составляет 13-15 %.

Адамантан является вполне доступным химическим соединением. Стоимость одного грамма у различных фирм-производителей не превышает одногодоллара США.
4. Физические свойства
4.1 Индивидуальное вещество
Химически чистый адамантан представляет собой бесцветное кристаллическое вещество, имеющее характерный камфорный запах. Он практически нерастворим в воде, но зато легко растворяется в неполярныхорганических растворителях.Адамантан имеет необычно высокую дляуглеводородовтемпературу плавления(268 °C), однако при этом медленносублимируетуже при комнатной температуре.Кроме того, он можетперегонятьсяс водяным паром.
4.2 Структурные свойства
Молекула адамантана включает в себя три конденсированныхциклогексановыхкольца, находящихся вконформации"кресло". Параметры молекулы адамантана были определены методом дифракции электронов ирентгеновских лучей. Было установлено, что длина каждой связи "углерод - углерод" составляет 1,54Å, а каждой связи "углерод - водород" - 1,112Å.

Молекула адамантана обладает высокой симметрией (точечная группаT d). Кристаллический адамантан существует в виде гранецентрированной кубической решётки (очень редкая для органических соединений пространственная группа
, a = 9,426 ± 0,008Å, четыре молекулы на ячейку).При охлаждении этой формы до температуры ниже −65 °C наблюдается фазовый переход с образованием объёмно-центрированной тетрагональной решётки (a = 6,641Å, c = 8,875Å).4.3 Спектральные свойства
Вспектре ЯМРадамантана присутствуют два слабо разрешённых сигнала, которые соответствуют протонам, находящимся около мостиковых и узловых атомов углерода. В спектре 1 H-ЯМР, записанном в CDCl 3 , сигналы протонов, расположенных возле узловых атомов углерода, наблюдаются при 1,873 м. д., а сигналы протонов при мостиковых атомах углерода - при 1,756 м. д. В спектре 13 С-ЯМР сигналы узловых и мостиковых атомов углерода проявляются при 28,46 и 37,85 м. д. соответственно.
Масс-спектрыадамантана и его производных довольно характеристичны. Положение основного пика в масс-спектре адамантана обусловлено наличием в продуктах ионизации иона
с соотношением m/z = 136. В результате фрагментации молекулярного иона обнаруживаются пики со значениями m/z равными 93, 80, 79, 67, 41, 39.Оптическая активность
Молекулы адамантана, содержащие четыре разных заместителя при узловых атомах углерода, являютсяхиральнымии оптически активными.При этом центр хиральности, как и у оптически активныхбифенилов, не приходится на какой-либо конкретный атом.R,S-номенклатурав этом случае может применяться так же легко.
трицикличесий мостиковый углеводород состава С 10 H 16 , молекула которого состоит из трех циклогексановых колец; пространственное расположение атомов углерода в молекуле адамантана то же, что и в кристаллической решетке алмаза. Согласно систематической номенклатуре, адамантан следует называть трицикло деканом.Обычно адамантан изображают одним из следующих способов:
В органической химии существует небольшое число веществ, которые вызывали громадный интерес химиков всего мира. Среди подобных соединений структуры бензола, ферроцена, карборана, фуллеренов и адамантана, есть и другие молекулярные структуры, вызывавшие и вызывающие интерес химиков-органиков. Вероятно, это во многом обусловлено необычностью структуры самих молекул, особенно высокой степенью их симметрии.
Строение адамантана. Углеродный скелет молекулы адамантана подобен структурной единице алмаза.
Именно поэтому название «адамантан» произошло от греческого «
adamas » алмаз. Экспериментально полученные структурные характеристики адамантана:
Подобная структура сохраняется практически у всех производных адамантана, что обусловлено высокой устойчивостью адамантанового каркаса. Адамантан родоначальник гомологического ряда семейства углеводородов алмазоподобного строения диамантана, триамантана и т.д.:
 .
.
На основе химии адамантана возникла и развилась одна из областей современной органической химии химия органических полиэдранов.
Адамантан, несмотря на небольшую молекулярную массу, обладает необычно высокой для насыщенных углеводородов температурой плавления 269° С. Эта аномально высокая температура обусловлена высокой симметрией жесткой алмазоподобной молекулы адамантана. Вместе с тем, относительно слабое межмолекулярное взаимодействие в кристаллической решетке приводит к тому, что углеводород легко возгоняется, частично уже при комнатной температуре.
В отличие от самого адамантана, его алкилзамещенные плавятся при гораздо более низких температурах (1-метиладамантан при 103° С, а 1-этиладамантан при 58° С) вследствие нарушения симметрии молекулы и увеличения колебательной и вращательной подвижности ее звеньев.
Несмотря на отсутствие в адамантане асимметрического атома углерода (атома углерода, связанного с четырьмя различными заместителями, расположенными в вершинах тетраэдра), производные адамантана, содержащие четыре различных заместителя в узловых положениях, оптически активны. Центр молекулы таких производных адамантана выполняет роль гипотетического асимметрического атома углерода.
Например:
 .
.
В данном случае оптическая активность обусловлена возникновением особого типа асимметрии асимметрии молекулярного тетраэдра. Величина оптического вращения для таких соединений невелика и редко превышает 1°.
Помимо оптической, для замещенных адамантанов характерна структурная изомерия , в зависимости от того к узловому или к мостиковому атому углерода присоединен заместитель. Например, возможны 1- и 2- пропиладамантаны, соответственно:

Для дизамещенных производных адамантана с одним мостиковым заместителем пространственная ориентация этого заместителя может быть аксиальной (а ) или экваториальной (е ), в зависимости от расположения заместителя по отношению к плоскости общего для обоих заместителей циклогексанового кольца (на рисунке выделено жирным) или ее можно обозначить как цис- и транс-. Например, для 1,3-дибромадамантана, возможны два изомера: 1,3 а -дибромадамантан и 1,3 е -дибромадамантан, соответственно:
 Получение адамантана и его алкилпрозводных
Единственным природным продуктом, содержащим адамантан и его гомологи, является нефть. Впервые адамантан был получен при исследовании нефти Годонинского месторождения (бывшая Чехословакия) в 1933 С.Ландой и В.Махачеком. Однако вследствие низкого содержания адамантана в нефти (оно обычно не превышает 0,001% масс.) получение его из этого сырья нецелесообразно. Количество адамантана в различных видах нефти зависит от ее химической природы нефти. Наиболее высое содержание адамантана в нефти нафтенового типа. Напротив, парафинистая нефть содержит адамантан в значительно меньших количествах. В нефти есть и алкильные производные адамантана, в частности, 1-метил, 2-метиладамантаны и 1-этиладамантан.
Получение адамантана и его алкилпрозводных
Единственным природным продуктом, содержащим адамантан и его гомологи, является нефть. Впервые адамантан был получен при исследовании нефти Годонинского месторождения (бывшая Чехословакия) в 1933 С.Ландой и В.Махачеком. Однако вследствие низкого содержания адамантана в нефти (оно обычно не превышает 0,001% масс.) получение его из этого сырья нецелесообразно. Количество адамантана в различных видах нефти зависит от ее химической природы нефти. Наиболее высое содержание адамантана в нефти нафтенового типа. Напротив, парафинистая нефть содержит адамантан в значительно меньших количествах. В нефти есть и алкильные производные адамантана, в частности, 1-метил, 2-метиладамантаны и 1-этиладамантан.
Так как выделение адамантана из нефти осложнено его низким содержанием, были разработаны методы химического синтеза этого вещества.
Впервые адамантан синтетическим путем удалось получить в 1941 швейцарцу Прелогу по следующей схеме:
 .
.
Однако общий выход адамантана составил всего 1,5 %. Были предложены улучшенные варианты вышеприведенного синтеза, но сложность синтеза, а также практическая невозможность синтеза замещенных адамантанов, ограничивает препаративную ценность этого метода.
Промышленно пригодный метод синтеза адамантана из легко доступного сырья был предложен и осуществлен Шлейером в 1957. Метод заключается в каталитической изомеризации трициклического углеводорода (по систематической номенклатуре трицикло декана) в адамантан:
 .
.
Метод практически интересен, поскольку циклопентадиен вполне доступное вещество, (его получают при крекинге нефтяных фракций как побочный продукт) и легко димеризуется. В зависимости от применяемого катализатора выходы адамантана изменяются в широком интервале. В качестве катализаторов могут применяться различные сильные кислоты Льюиса , такие как
AlCl 3 , SbF 5 . Выходы колеблются от 15 до 40%.Этот метод пригоден и для синтетического получения различных алкил замещенных адамантанов:
 .
.
Характерно, что наличие алкильных групп существенно увеличивает выход конечных продуктов изомеризации.
Высокий выход алкиладамантанов получают при изомеризации (над галогенидами алюминия или комплексами на их основе) трициклических пергидроароматических углеводородов состава С 12 С 14: пергидроаценафтена, пергидрофлуорена, пергидроантрацена и других углеводородов.

Выход в последней реакции составляет 96%.
Доступность исходных соединений (соответствующие ароматические углеводороды в значительных количествах легко выделяются из жидких продуктов коксования каменного угля), высокий выход конечных продуктов изомеризации делают этот метод промышленно привлекательным.
В описанных методах жидкофазной каталитической изомеризации применяются катализаторы (
AlCl 3 , SbF 5), которые обладают рядом существенных недостатков: повышенная коррозионная активность, нестабильность, невозможность регенерации, образование в процессе реакции значительных количеств смолы. Это послужило поводом для исследования изомерных превращений полициклоалканов с использованием стабильных гетерогенных катализаторов кислотного типа, получаемых на основе оксидов металлов. Были предложены катализаторы на базе оксида алюминия, которые позволяют получать алкиладамантаны с выходами до 70%.Каталитические методы изомеризации полициклоалканов эффективные методы получения углеводородов ряда адамантана, многие из них имеют препаративное значение, а процесс получения адамантана изомеризацией гидрированного димера циклопентадиена реализуется в промышленном масштабе.
Однако по мере повышения молекулярной массы и увеличения числа циклов в исходном углеводороде скорость перегруппировки в адамантаноидные углеводороды замедляется. В ряде случаев методы изомеризации не дают желаемого результата. Так, с их помощью нельзя получить 2-замещенные алкил- и ариладамантаны, кроме того, продукты реакции, как правило, состоят из смеси нескольких изомеров, и нужно их разделить, поэтому широкое развитие приобрели синтетические методы получения углеводородов ряда адамантана, основанные на применении функциональных производных адамантана в качестве исходных веществ, а также методы циклизации построения структуры адамантана, исходя из алифатических моно- и бициклических соединений. Синтезы на основе функциональных производных широко используются для получения индивидуальных алкил-, циклоалкил-, и ариладамантанов. Методы циклизации используют обычно при синтезах полифункциональных производных адамантана, углеводородов адамантаноидного строения и их производных.
Одним из первых успешных синтезов 1-метиладамантана был многостадийный синтез на основе 1-бромадамантана (обычно адамантильный радикал обозначается в схемах реакций как
Ad ): .
.
Позднее были найдены и другие более эффективные пути синтеза 1-метиладамантана.
Метод, приведенный ниже можно рассматривать как общий метод синтеза алкиладамантанов, полизамещенных в узловых положениях. Он позволяет путем постепенного наращивания углеводородной цепи получить алкиладамантаны с различной длиной алкильных групп нормального строения.

Прямой синтез производных адамантана, замещенных в мостиковые положения, затруднен вследствие низкой реакционной способности мостиковых атомов углерода адамантанового ядра. Для синтеза 2-алкилпроизводных адамантана используют взаимодействие реактивов Гриньяра или алкиллитиевых производных с легко доступным адамантаноном. Так, 2-метиладамантан может быть получен по схеме:
 .
.
Что касается других методов получения адамантановых структур, то наиболее распространенными являются методы синтеза циклизацией производных бициклононана. Хотя такие методы многостадийны, они позволяют получать производные адамантана с заместителями, которые трудно синтезировать иным путем:


.
Адамантильный катион генерируется и из 1-хлор-, оксиадамантанов в суперкислотах (SbF 5) или в «магической кислоте» (SbF 5 в HSO 3 F) в среде SO 2 и SO 2 ClF.
Наиболее употребительные ионные реакции, протекающие в узловых положениях адамантанового ядра:
Адамантан и его производные бромируются обычно молекулярным бромом в жидкой фазе, это ионный процесс, катализируемый кислотой Льюиса и нечувствительный к инициаторам радикального типа. Применение катализаторов Фриделя
Крафтса позволяет заместить все четыре атома водорода в узловых положениях адамантанового ядра на бром: .
.
В условиях ионного галогенирования процесс идет селективно по узловым атомам углерода адамантанового ядра.
В отличие от ионного галогенирования, свободнорадикальное галогенирование самого адамантана и его производных приводит к смеси продуктов, состоящих из 1- и 2-замещенных производных.
Для получения фторпроизводных адамантана используют 1-адамантанол:
 .
.
Галогензамещенные адамантана широко используются для синтеза других функционально замещенных адамантанов. Реакционная способность галогенпроизводных адамантана больше, чем у других насыщенных углеводородов. Окисление адамантана серной кислотой представляет собой важный препаративный метод, поскольку позволяет получать адамантанон с высоким выходом:
 .
.
В тоже время взаимодействие адамантана с концентрированной серной кислотой в среде трифторуксусного ангидрида позволяет получить смесь 1- и 2-адамантанолов, с преимущественным содержанием первого из них:
 .
.
Для синтеза карбоновых кислот адамантанового ряда чаще всего используют реакцию карбоксилирования. Кох и Хааф впервые в 1960 осуществили таким путем прямой синтез 1-адамантанкарбоновой кислоты. Реакцию проводят в среде концентрированной серной кислоты или олеума, что обеспечивает образование адамантилкатионов.

 .
.
Удобнее получать 1-аминоадамантан можно получать по одностадийной реакции Риттера, заключающейся во взаимодействии самого адамантана или 1-бромадамантана с нитрилом (обычно ацетонитрил) в присутствии трет -бутилового спирта под действием брома в серной кислоте:
 .
.
Последующий гидролиз образовавшегося амида приводит к 1-аминоадамантану.
Среди реакций функционализации адамантана есть интересный способ активации С-H -связи в адамантановом ядре, предложенный Олой с помощью хлористого алюминия в хлористом метилене в присутствии трехлористого фосфора. В результате реакции образуются дихлорфосфорилированные производные с выходами 4060%.

Несмотря на необычность структуры адамантана, реакции в которые он вступает, довольно традиционны для органической химии. Особенность же адамантана проявляется за счет либо стерических эффектов, связанных с большим размером адамантильного радикала, либо с возможностью образования относительно устойчивого адамантильного катиона.
Применение. Перспективность применения производных адамантана обуславливается набором специфических свойств: относительно большой размер адамантильного радикала (его диаметр составляет 5 Å), высокая липофильность (растворимость в неполярных растворителях), конформационная жесткость. Последние два свойства особенно важны при создании новых лекарственных препаратов. Введение адамантильного радикала повышает, в целом, термическую стабильность вещества и его стойкость к окислению и радиационному облучению, что важно, в частности, при получении полимеров со специфическими свойствами.Все это стимулировало широкомасштабный поиск новых лекарственных препаратов, полимерных материалов, присадок к топливам и маслам, взрывчатых веществ, жидких ракетных топлив, неподвижных фаз для газожидкостной хроматографии на основе производных адамантана.
Сейчас сам адамантан не используется, но ряд его производных находит широкое применение.
В наибольшей степени производные адамантана применяются в фармацевтической практике.
Так, препараты ремантадин (гидрохлорид 1-(1-адамантил)этиламина), и адапромин (
a -пропил-1-адамантил-этиламина гидрохлорид) используются как лекарства при эффективной профилактике вирусных инфекций, а амантадин (гидрохлорид 1-аминоадамантана) и глудантан (глюкуронид 1-аминоадамантана) эффективны при паркинсонизме, порождаемом различными причинами, в частности, при нейролептическом и посттравматическом синдроме.Полимерные аналоги адамантана запатентованы как противовирусные соединения, в том числе, в отношении ВИЧ , полимерные аналоги адамантана.
Замещенные амиды адамантанкарбоновой кислоты могут служить снотворными средствами. Введение адамантильного остатка в 2-оксинафтохинон приводит к получению антималярийных препаратов. Адамантиламиноспирты и их соли обладают выраженным психостимулирующим действием и при этом мало токсичны. Некоторые
N -(адамант-2-ил)анилины проявляют нейротропную активность, а биологическая активность N -(адамант-2-ил)гексаметиленимина проявляется по отношению к паркинсоническому синдрому.Алкилпроизводные адамантана, в частности 1,3-диметиладамантан, используются как рабочие жидкости в некоторых гидравлических установках. Целесообразность такого их использования объясняется высокой термической стабильностью диалкилпроизводных, их низкой токсичностью и большой разницей между критической температурой и температурой кипения.
В химии высокомолекулярных соединений введение адамантильного заместителя позволило во многих случаях улучшить эксплуатационные характеристики полимерных материалов. Обычно, полимеры, содержащие в своем составе адамантильный фрагмент, термостойки и их температура размягчения достаточно высока. Они достаточно устойчивы и к гидролизу, окислению, фотолизу. По этим свойствам адамантансодержащие полимерные материалы превосходят многие известные промышленные полимеры и могут найти применение в различных областях техники в качестве конструкционных, электроизоляционных и иных материалов.
Владимир Корольков
ЛИТЕРАТУРА Багрий Е.И. Адамантаны: Получение, свойства, применение . М. , Наука, 1989Морозов И.С., Петров В.И., Сергеева С.А. Фармакология адамантанов . Волгоград: Волгоградская мед. академия, 2001
Введение
В медицине всё ещё стоит вопрос об эффективности лечения патологий системы свёртывания крови, в частности таких нарушений, как тромбоцитоз (патологическое увеличение числа тромбоцитов) и тромбоцитопения (уменьшение числа тромбоцитов в крови). Повышенная способность тромбоцитов к агрегации наблюдается при ряде заболеваний: ишемической болезни сердца, атеросклерозе, диабетических ангиопатиях и др., что значительно осложняет течение болезни . Путём направленного фармакологического воздействия можно снижать патологически возникшую гиперагрегацию тромбоцитов, или наоборот, повышать их способность к агрегации.
В данной работе исследовано влияние производных адамантана с различными характерами заместителей на индуцированную агрегацию тромбоцитов человека. Ранее было показано, что амиды N-[(адамантоил-1)-фенил]-антраниловой кислоты проявляли выраженное ингибирование серотонинового отёка, предполагалось, что это явление вызвано блокированием серотониновых рецепторов сосудистой стенки этими амидами .
Научная новизна работы: впервые исследованы вновь синтезированные производные адамантана, способные усиливать или снижать эффект известных индукторов агрегации тромбоцитов посредством модуляции функционирования соответствующих тромбоцитарных рецепторов.
Результаты работы имеют практическое значение – исследованные соединения после более детального изучения механизма их действия могут быть использованы в качестве медицинских препаратов.
Цель работы: изучить механизмы влияния вновь синтезированных производных адамантана на индуцированную агрегацию тромбоцитов человека.
Задачи исследования:
1. Выявить среди исследуемых соединений синергистов (или агонистов) и антагонистов серотонина;
2. Выяснить механизмы влияния этих соединений на индуцируемую агрегацию тромбоцитов человека;
3. Проверить наличие свойств агонистов агрегирующих тромбоциты веществ у всех исследуемых соединений;
4. Выявить синергистов и антагонистов серотонина;
5. Выявить синергистов и антагонистов АДФ;
6. Выявить синергистов и антагонистов адреналина.
Глава 1. Обзор литературы
1.1. Механизмы агрегации тромбоцитов человека
Тромбоциты – плоские безъядерные клетки крови неправильной округлой формы, образующиеся из мегакариоцитов костного мозга путём отщепления от них участков цитоплазмы. Образование тромбоцитов регулируется гликопротеиновым гормоном тромбопоэтином. Нормальные тромбоциты – клетки диаметром 1-5 мкм, толщиной 0,5-0,75 мкм. Содержание тромбоцитов в крови здорового человека составляет 180-320 тыс. в 1 мл. Молодые тромбоциты проходят период созревания – восемь суток, зрелые циркулируют в кровотоке от пяти до 11 суток, с последующим разрушением в печени и селезёнке. Число тромбоцитов в крови относительно постоянно, но ночью оно снижается, а возрастает при пищеварении, тяжёлой мышечной работе, беременности .
На мембране тромбоцитов адсорбируются факторы свёртывающей системы крови. Тромбоциты способны выделять из фосфолипидов своих клеточных мембран арахидоновую кислоту и превращать её в тромбоксаны (при участии тромбоксансинтетазы), которые повышают агрегационную активность тромбоцитов. Примыкающая к оболочке тромбоцита область неструктурированной цитоплазмы – гиаломер. В гиаломере после активации тромбоцита при электронной микроскопии становятся видны микрофиламенты из актина, миозина и тропомиозина, составляющие вместе с микротрубочками цитоскелет клетки, который определяет её форму и способность перемещать органеллы, прикрепляться к поверхностям и образовывать псевдоподии . Актиновые филаменты имеют диаметр 3-5 нм, а миозиновые 6-10 нм. Микрофиламенты контактируют между собой, плазматической мембраной и мембранами гранул . Микротрубочки в основном состоят из тубулина, их диаметр 25-30 нм. Концентрическая система из 5-30 микротрубочек тромбоцитов образует периферический пучок под плазматической мембраной вдоль экватора клетки. Также присутствуют и другие немембранные структуры – гранулы гликогена.
В тромбоцитах различают две системы мембран: плотную тубулярную систему (ПТС) и систему открытых каналов. ПТС – система узких трубочек диаметром 50 нм, подобная ЭПР скелетной мышцы. Эти трубочки не контактируют с системой открытых каналов, органеллами и плазматической мембраной, расположены и по экватору клетки и среди органелл . Система открытых каналов сообщается с гранулами тромбоцитов и с плазматической мембраной, за счёт чего и осуществляется секреция тромбоцитами специфических соединений в процессе реакции высвобождения. Поверхность тромбоцитов может увеличиваться за счёт системы открытых каналов . Центральная часть тромбоцитов (грануломер) содержит органеллы и множество гранул различной структуры, формы и величины («электроноплотные» гранулы, α-гранулы, лизосомы). В электроноплотных гранулах содержатся АДФ, АТФ, ГТФ, ГДФ, неорганический фосфат, серотонин, Са 2+ ; в α-гранулах – трансформирующий ростовой фактор, β-тромбоглобулин, антигепарин (фактор пластинок 4), фибронектин, альбумин, фибриноген, проакцелерин (фактор V), антиген фактора Виллебранда (фактор VIII), калликреин, α 2 -антиплазмин, тромбоспондин, гистамин; в лизосомах – кислые гидролазы (β-гексозаминидаза, β-галактозидаза, β-глюкуронидаза, β-арабинозидаза, β-глицерофосфатаза, арилсульфотаза) . В тромбоцитах имеются гликолитические ферменты и ферменты пентозофосфатного цикла, цикла лимонной кислоты и дыхательной цепи, АТФаза .
Важнейшим свойством тромбоцитов является способность к агрегации, в процессе которой в них синтезируется, а затем секретируется ряд белков и биологически активных веществ, стимулирующих свёртывание крови (гемостаз). Тромбоциты принимают участие и в защите организма от чужеродных агентов. Они обладают фагоцитарной активностью, содержат IgG, являются источником лизоцима и β-лизинов, способных разрушать мембрану некоторых бактерий.
Гемостаз осуществляется в 4 фазы: сокращение сосуда, образование тромбоцитарной пробки, формирование красного тромба и его ретракция, полное или частичное растворение тромба . Различают внешний путь свёртывания (в месте повреждения сосудов, индуцируется коллагеном) и внутренний путь свёртывания (в области замедленного кровотока на патологически изменённой поверхности сосудистой стенки, индуцируется факторами плазмы), они различаются начальными этапами, объединяясь на стадии активации фактора Х. В настоящее время известно много факторов свёртывания крови, отсутствие любого из которых может привести к нарушению процесса коагуляции. В основном они представляют собой протеолитические ферменты (факторы XII, XI, X, IX, VII, II и калликреин – сериновые протеазы), присутствующие в крови в неактивной форме в виде проферментов. В процессе свёртывания они активируют друг друга в каскадной последовательности реакций. Активированные факторы обозначаются прибавлением буквы «а», например, X а. (рис. 1)
Рис. 1. Схема свёртывания крови и фибринолиза. ФП 3 – тромбоцитарный фактор 3 (по Циммерману, 1996)
Сразу после травмы наблюдается первичный спазм кровеносных сосудов, благодаря чему кровотечение в первые секунды может не возникнуть или носит ограниченный характер. Первичный спазм сосудов обусловлен выбросом в кровь в ответ на болевое раздражение адреналина и норадреналина и длится 10-15 с. Повреждение сосудов сопровождается немедленной активацией тромбоцитов вследствие появления высоких концентраций АДФ (из травмированных клеток сосудов), а так же обнажением субэндотелия, коллагеновых и фибриллярных структур . В дальнейшем наступает вторичный спазм, обусловленный активацией тромбоцитов и выбросом в кровь сосудосуживающих агентов – серотонина, тромбоксана А 2 , адреналина и др. (реакция высвобождения).
Просвет повреждённых сосудов уменьшается и перекрывается массой тромбоцитов, прилипших к коллагеновым волокнам. Тромбоксан А 2 и АДФ, выделяющиеся при разрушении внутриклеточной структуры активированных тромбоцитов, вызывают присоединение новых тромбоцитов. АДФ также может высвобождаться из разрушенных эритроцитов , и способствует превращению кровяных пластинок в диски, а также вызывает секрецию тромбоцитарных белков из электроноплотных и α-гранул . Тромбоциты распластываются, у них появляется до 10 «отростков», которые могут в 5-10 раз превышать диаметр самой клетки . Организованное кольцо микротрубочек распадается, изменяется распределение микрофиламентов и микротрубочек, микротрубочки могут образовывать клубочки .
На нескольких из этапов гемокоагуляции существуют перекрёстные взаимодействия между внешней и внутренней системами свёртывания, благодаря чему обеспечиваются «альтернативные» пути для процессов коагуляции .
Агрегация тромбоцитов наступает одновременно с их адгезией, это активный процесс, протекающий с затратой энергии. Адгезия тромбоцитов к волокнам соединительной ткани по краям раны обусловлена олигомерным гликопротеином, содержащимся в субэндотелии и кровяных пластинках, – фактором Виллебранда. Он образует мостики между субэндотелиальными структурами и специфическими рецепторами (гликопротеин Ib) на мембране тромбоцитов . Фибриноген – белок, содержащийся в плазме и тромбоцитах, тоже способный образовывать между тромбоцитами связующие мостики, что приводит к появлению сети фибрина и тромбоцитарной пробки. Частичный протеолиз молекулы фибриногена наступает под действием тромбина. Для образования ковалентных связей необходим фибринстабилизирующий фактор XIII (трансглутаминаза), активируемая тромбином в присутствии ионов кальция. Трёхмерная сеть волокон фибрина удерживает клетки крови, формируется красный тромб (рис. 1). Благодаря контрактильному белку тромбостенину тромбоциты подтягиваются друг к другу, тромбоцитарная пробка сокращается и уплотняется, т. е. наступает её ретракция . Сгусток становится более плотным и стягивает края раны, что облегчает её зарастание клетками соединительной ткани. В норме остановка кровотечения из мелких сосудов занимает от двух до четырёх минут .
Таким образом, при повреждении кровеносного сосуда в крови появляются вещества, инициирующие процесс агрегации тромбоцитов. Характер действия этих веществ разный, так, слабыми агрегирующими агентами являются АДФ, серотонин, адреналин, вазопрессин, – они действуют первыми, лишь запуская каскад последующих реакций. Более быстро агрегация происходит под действием коллагена, тромбина, ионофора А23187. Агрегация тромбоцитов может носить обратимый характер (вслед за агрегацией наступает дезагрегация, распад агрегатов), что зависит от недостаточной дозы агрегирующего агента или от наличия в крови простациклина PGI 2 , синтезирующегося в эндотелии сосудов, который ингибирует агрегацию тромбоцитов . Травма сосуда всегда сопровождается образованием сразу нескольких индукторов агрегации, способных инициировать агрегацию тромбоцитов разными путями, и их концентрация достаточно высока, поэтому дезагрегация, как правило, не случается.
Биохимические механизмы активации тромбоцитов с их последующей агрегацией различны, что обусловлено различным характером инициаторов агрегации. Однако, все известные агреганты так или иначе способствуют увеличению концентрации Са 2+ в цитоплазме тромбоцита.
Общее содержание Са 2+ в тромбоцитах составляет около 60 нмоль/10 9 клеток. Четверть этого кальция связана с мембранами тромбоцитов, но значительное его количество содержится в тубулярной системе и плотных гранулах. Концентрация свободных ионов Са 2+ в цитоплазме тромбоцитов (10 -7 моль/л)почти в 1000 раз ниже, чем в плазме (10 -3 моль/л). Такая разница в концентрации этих ионов по обе стороны плазматической мембраны поддерживается мембранными Са 2+ -АТФазами, которые используют энергию гидролиза АТФ для откачивания Са 2+ из цитоплазмы в окружённые мембранами клеточные органеллы, а так же аденилатциклазами и фосфодиэстеразами, контролирующими уровень цАМФ. Активированная аденилатциклаза катализирует образование цАМФ из АТФ, фосфодиэстераза осуществляет гидролиз цАМФ до 5-АМФ. Аденилатциклаза тромбоцитов локализована на внутренней стороне плазматической мембраны, на мембранах плотной тубулярной системы и системы открытых каналов. Фосфодиэстераза находится в цитоплазме. Активация аденилатциклазы происходит посредством взаимодействия простагландинов Е 1 , I 2 и D 2 со специфическими рецепторами мембран, сопряжёнными с ГТФ-связывающими белками (G-белки). Для этих рецепторов цАМФ служит внутриклеточным медиатором. Активированные рецепторы действуют на G-белки, которые стимулируют аденилатциклазу, образуется цАМФ, активируются цАМФ-зависимые ферменты (протеинкиназы), катализирующие фосфорилирование небольшой группы тромбоцитарных белков .
Адреналин, связываясь со специфическими рецепторами мембраны тромбоцитов, посредством G-белков стимулирует образование тромбоксана А 2 . Предполагается, что ускоряющий агрегацию тромбоцитов эффект адреналина связан с модуляцией мембран при его взаимодействии с α-адренорецепторами и изменением её проницаемости к ионам Са 2+ . Показано, что антагонисты α 2 -адренорецепторов способны блокировать агрегацию, а агонисты усиливают её, поскольку активированные G i -белки ингибируют аденилатциклазу . Через β-адренорецепторы происходит замедление агрегации тромбоцитов. Стимуляция β-адренорецепторов посредством G s -белков приводит к активации аденилатциклазы, т. е. катехоламины способны регулировать активность тромбоцитов, влияя на уровень цАМФ в клетке. Адреналин вызывает агрегацию тромбоцитов без изменения их формы . Адреналин-индуцированное изменение оптической плотности проб наблюдается через 30 с после его добавления .
Активация тромбоцитов тромбином, коллагеном, фактором активации тромбоцитов (ФАТ) и некоторыми другими агрегантами происходит через образование из фосфолипидов плазматической мембраны (фосфатидилхолина и фосфатидилинозитола) арахидоновой кислоты . Активированная через сопряжённые с G s -белками рецепторы для ФАТ фосфолипаза С катализирует этот процесс. Арахидоновая кислота способна метаболизировать, вновь включаться в фосфолипиды или диффундировать из тромбоцитов. Высвобождение арахидоновой кислоты из фосфолипидов тромбоцитов могут катализировать также фосфолипаза А 2 и глицеролипаза. Образовавшаяся арахидоновая кислота может окисляться двумя ферментами: цитоплазматической циклооксигеназой (ЦОГ) и мембранносвязанной липоксигеназой (ЛОГ). При действии ЦОГ образуются нестабильные циклические эндопероксиды, которые превращаются в простагландины G 2 и Н 2 , тромбоксан А 2 , простациклин с дальнейшим образованием из них 6-кето-простагландина Е 1 , 12-гидрокси-5,8,10-гептадекатриеновой кислоты, тромбоксана В 2 и малондиальдегида. Конечными продуктами липоксигеназного пути окисления арахидоновой кислоты являются 12-гидрокси-5,8,10,14-эйкозатетраеновая кислота (12-НЕТЕ) и лейкотриены . Образующиеся из арахидоновой кислоты простагландины и тромбоксан А 2 (ионофор кальция) являются агрегантами. Простациклин, синтезируемый клетками эндотелия кровеносных сосудов, напротив, мощный ингибитор агрегации тромбоцитов.
Связывание АДФ со специфическими рецепторами плазматической мембраны тромбоцита приводит к увеличению концентрации ионов кальция в цитоплазме клеток (увеличивается его поступление из внешней среды и из внутриклеточных запасов) . Кроме того, происходит активация фосфолипазы А 2 , которая стимулирует высвобождение арахидоновой кислоты из фосфолипидов мембраны и образование из неё тромбоксана А 2 – активного проагреганта и вазоконстриктора. Форма тромбоцитов изменяется без сопутствующей активации фосфолипазы С, что сопровождается ингибированием аденилатциклазы . Реализация метаболических эффектов активации АДФ-рецепторов тромбоцитов осуществляется через сопряжение с мембранным G-белком. Сегодня известно, что при активации на поверхности каждого тромбоцита появляется до 50–100 тысяч гликопротеиновых рецепторов . Экспериментальные данные многих клинических исследований свидетельствуют, что удаление из среды АДФ или связывание АДФ-рецепторов с конкурентным антагонистом существенно угнетает агрегацию тромбоцитов .
Для серотонина на поверхности тромбоцитов существует два типа рецепторов – ионные каналы и рецепторы, сопряжённые с G-белком . Согласно , механизм действия серотонина состоит в регуляции цАМФ-зависимых путей, и поэтому реакция на серотонин способна блокироваться стимуляторами аденилатциклазы и ингибиторами фосфодиэстеразы. Серотонин стимулирует поступление ионов кальция из плазмы в тромбоциты через рецепторуправляемые каналы для двухвалентных катионов, запуская фосфатидилинозитоловый цикл с образованием фосфатидной кислоты, являющейся кальциевым ионофором. Активация G-белков, сопряжённых с другим типом серотониновых рецепторов приводит к активации фосфолипазы С, которая гидролизует трифосфатидилинозитол (или фосфатидил-4,5-бифосфат) в дифосфатидилинозитол (фосфатидилинозитол-4-фосфат) с образованием 1,2-диацилглицерола (ДАГ), который фосфорилируется в фосфатидную кислоту (ФК). Это самые ранние этапы активации тромбоцитов. Диацилглицерол стимулирует Са 2+ -активируемые фосфолипидзависимые протеинкиназы (в частности, ПК С), которые фосфорилируют белки. Фосфатидная кислота – кальциевый ионофор, высвобождает кальций из внутриклеточных депо, присоединяя к своим фосфатным группам. Ионы кальция стимулируют работу актина и миозина микрофиламентов, в результате чего гранулы тромбоцитов перемещаются к системе открытых каналов, осуществляется реакция высвобождения собственных агрегантов, что усиливает собственный ответ клетки и активирует другие тромбоциты.
Таким образом, активация тромбоцитов начинается при взаимодействии агрегантов с рецепторами плазматической мембраны. Важную роль в передаче сигнала внутрь тромбоцита играют G-белки плазматической мембраны, а также изменение проницаемости мембраны для ионов (если рецептор является ионофором) .
1.2. Роль рецепторов плазматической мембраны в процессах агрегации тромбоцитов человека
В настоящее время установлено, что важным моментом трансмембранной передачи сигналов является изменение интенсивности транспорта и внутриклеточной концентрации различных ионов. Одной из основных систем, приводящих к изменению внутриклеточной концентрации ионов, являются селективные ионные каналы биомембран, представляющие собой интегральные мембранные белки, способные при определённых внешних воздействиях (изменение потенциала на мембране, действие медиатора или гормона) избирательно менять проницаемость мембраны для конкретного вида ионов . Агрегацию тромбоцитов и вазоконстрикцию индуцирует 5-гидрокситриптамин (серотонин), который также синергически усиливает эффекты тромбоксана А 2 , ангиотензина II, АДФ и коллагена . Эта активность проявляется через взаимодействие со специфическими рецепторами на поверхности тромбоцитов – 5-НТ-рецепторами (рецепторы 5-гидрокситриптамина) . Серотонин (5-гидрокситриптамин) является производным триптофана и синтезируется в клетках пищеварительного тракта, нервных (серотонинэргических) и тучных клетках; способен накапливаться в специальных гранулах тромбоцитов. Физиологическое действие серотонина в организме человека разнообразно, он является лигандом для рецепторов клеток практически всех внутренних органов и тканей . Фармакологические эффекты серотонина коррелируют главным образом с возбуждением 5-НТ 2 рецепторов . Исследователями синтезировано множество соединений, способных связываться с этими рецепторами и модулировать их функционирование. Установлено, что рецепторы для 5-гидрокситриптамина (серотонина) находятся на клетках практически всех внутренних органов человека, регулируя многие процессы жизнедеятельности. Классификация 5-НТ рецепторов на семь семейств (от 5-НТ 1 до 5-НТ 7) основана на структурных, функциональных и фармакологических критериях . Семейства в свою очередь делятся на подсемейства . Выяснено, что некоторые 5-НТ рецепторы связаны с G-белками . Рецепторы серотонина, расположенные на тромбоцитах и клетках гладкой мускулатуры сосудов относятся к подтипу 5-НТ 2А рецепторов, остальные представители этого подтипа – 5-НТ 2В и 5-НТ 2С локализованы только на нейронах .
Есть данные о примерном расположении в мембране и структуре активного центра 5-НТ 4 рецептора.
Представители некоторых семейств 5-НТ рецепторов синтезируются с использованием методов генной инженерии invitro, выделяются методом микродиализа или экстракцией из клеточных мембран . На плазматической мембране тромбоцитов локализованы два типа рецепторов для серотонина .
Исследованиями показана зависимость снижения агрегационной способности тромбоцитов от геометрии молекулы изучаемого вещества. Есть данные о том, что сродство к 5-НТ 2 рецепторам увеличивается при наличии объёмного адамантил-заместителя . Так же доказано антагонистическое действие SB-699551-A (3-циклопентил-N--N-[(4¢-{[(2-фенилэтил)амино]метил}-4-бифенилил)метил]пропанамид дигидрохлорид), имеющего объёмное, сложное молекулярное строение на 5-НТ 5А рецепторы мозга свиньи .
Ионотропные рецепторы для серотонина являются членами суперсемейства мембранных белков, которое включает также рецепторы для никотина, глицина, γ-аминомасляной кислоты (ГАМК). Все они обладают одинаковыми свойствами – один и тот же белок является и рецептором и ионным каналом, сформированным различными комбинациями 2α, β, γ, δ и ε субъединиц. Лиганд рецептора этого суперсемейства связывается с α-субъединицей и стимулирует открытие неспецифического ионного канала .
Большинство рецепторов относятся к семейству мономерных гликопротеинов, семикратно пересекающих мембрану. Они выполняют разнообразные биологические сигнальные функции. К таким рецепторам относятся адренорецепторы и рецепторы для АДФ тромбоцитов, а так же часть 5-НТ рецепторов тромбоцитов. АДФ активирует тромбоциты через их три аденозинфосфат-зависимых рецептора: Р2Х, П2И и Р2Т . Гликопротеиновые рецепторы имеют очень древнее происхождение. Их используют, например, клетки дрожжей, которые выделяют необходимые для спаривания полипептидные факторы и распознают их с помощью поверхностных рецепторов, представляющих собой все те же семикратно пересекающие мембрану рецепторы.
Уникальная структура лиганд-связывающих участков семикратно пересекающих мембрану рецепторов позволяет связывать лиганды различной природы и молекулярной массы в широком диапазоне от 32 для Ca 2+ до более чем 10 2 кДа для гликопротеинов.
Большинство обычных низкомолекулярных гормонов (типа адреналина и ацетилхолина) связывается с участками внутри гидрофобного ядра (a). Пептидные и белковые лиганды присоединяются к внешней поверхности рецептора (b, c). Некоторые лиганды низкого молекулярного веса, Ca 2+ и аминокислоты (глутамат, ГАМК) связываются с длинными участками на N-конце, индуцируя их переход в новую конформацию, в которой длинный участок взаимодействует с рецептором (d). В случае рецепторов, активируемых отрезающей протеазой (e), новый N-конец действует как автолиганд. Отрезанный пептид может также взаимодействовать с другим рецептором.
1.3. Биологическая активность производных адамантана
Производные адамантана как физиологически активные вещества находят широкое применение с 70-х годов XX века . Сам адамантан (трициклодекан, С 10 Н 16) принадлежит к числу трициклических нафтенов мостикового типа (рис. 6).
Рис. 6. Структура молекулы адамантана.
Молекула его состоит из трёх конденсированных циклогексановых колец в конформации кресла. Пространственная модель молекулы адамантана – высокосимметричная конструкция с малой поверхностью и незначительными силами межмолекулярного взаимодействия в кристаллической решётке . Из всех трициклических углеводородов адамантан наиболее устойчив, что объясняется тетраэдрической направленностью связей всех атомов углерода и их фиксированным положением.
Биологическая активность производных адамантана обусловлена симметрией и объёмностью пространственного строения, значительной
липофильностью жёсткого углеводородного каркаса адамантана, что позволяет им легко проникать через биологические мембраны . Поэтому модификация органических соединений с помощью адамантильного радикала значительно изменяет их биологическую активность, нередко усиливая её. С помощью метода спиновых меток было показано, что адамантан, попадая в липидный бислой, способен разрушать гексагональную упаковку метиленовых группировок, характерную для двойного слоя фосфолипидов, и нарушать осевое расположение алкильных цепей фосфолипидов, модифицируя тем самым функциональные свойства клеточных мембран . Принимая во внимание важность порядка расположения метиленовых групп липидов биологических мембран как фактора функционирования связанных с мембранами ферментов, можно отметить косвенное влияние адамантана на их активность.
На данный момент синтезировано более 1000 новых производных адамантана. Фармакологическое изучение показало наличие среди них веществ, обладающих выраженной психотропной, иммунотропной, противовирусной, курареподобной, антикаталептической, противоаллергической активностями, а также соединений, влияющих на ферментативную систему печени . Амиды адамантанкарбоновых кислот проявляют антибактериальную активность .
Есть данные о результатах модификации молекулы энкефалина аминокислотами ряда адамантана. Введённый в положение 5 молекулы энкефалина (S)-адамантилаланин придаёт опиоидному пептиду резистентность по отношению к ферментам, легко разрушающим немодифицированный энкефалин (химотрипсин, проназа, нейтральная протеаза, термолизин) .
Показано, что физиологической активностью обладают азотсодержащие производные. Первым в медицинскую практику в 1966 г. вошёл гидрохлорид 1-аминоадамантана, обладающий противовирусной активностью в отношении штаммов вирусов типа А 2 , его фирменные названия: мидантан, симметрел, амантадин. Эти препараты используются для профилактики респираторных заболеваний, т. к. обладают способностью блокировать проникновение вируса в клетку. Предполагают, что данные препараты способны работать на начальных этапах репродукции вируса, блокируя синтез вирусоспецифичных РНК . Противовирусная активность некоторых аминопроизводных адамантана связана с их способностью ингибировать ПК С . Ремантадин (полирем, флумадин), как липофильное слабое основание способен повышать рН эндосомального содержимого и препятствовать депротеинизации вируса .
В клинической практике для лечения вирусных заболеваний применяются также такие препараты, как ацикловир (виролекс, герпесин, зовиракс, лизавир, суправиран), диданозин, фоскарнет (триаптен), ганцикловир (цимевен), ламивудин, рибавирин (виразол, рибамидил), ставудин, трифлуридин, видарабин, зальцитабин (хивид), зидовудин (азидотимидин, ретровир) . Однако, у большинства этих препаратов относительно узкий спектр противовирусного действия, их недостатком является наличие разнообразных побочных реакций, появление резистентных штаммов вирусов и др. .
Противовирусной активностью в отношении штаммов вирусов типа А 2 обладают и производные алкиладамантанов: 1-гидрокси-3,5диметил-7-этиладамантан, 1-метокси-3,5диметиладамантан, которые, в отличие от мидантана, показали высокую противовирусную активность и в отношении штаммов strainsofrhinoviruses и herpessimplex . Ряд гидрокси-, галоген- и меркаптопроизводных амидов адамантана тоже обладают противовирусной активностью .
Показано, что амантадин способен препятствовать развитию очагов саркомы на зародышевой культуре, другие производные адамантана могут служить снотворными средствами, антималярийными препаратами, инсектицидами . В экспериментах с использованием ВИЧ-инфицированных человеческих лимфобластоидных клеток показано, что некоторые производные адамантана обладают анти-ВИЧ-активностью . Мидантан используется в неврологической клинике для лечения болезни Паркинсона и паркинсонического синдрома. Подобную активность проявляют хлорангидриды 3,5,7-алкилзамещённых 1-аминоадамантанов, некоторые из которых обладают антагонистическими дофамину свойствами. Некоторые четвертичные аммониевые основания с 2-адамантильным радикалом способны действовать как миорелаксанты периферического действия (курареподобная активность). Производные 1-аминоадамантана и 3,3-диамино-1,1-диадамантила противокаталептически активны, бактериостатическое влияние оказывают адамантанкарбоновые кислоты и фосфаты адамантантиолов и их производных. Диалкиламиновые эфиры адамантантиокарбоновой кислоты проявляют бактерицидную, фунгицидную, гербицидную активность . Натриевая соль β-(1-адамантан)-пропионовой кислоты обладает желчегонным действием. 1-адамантиламмоний-β-хлорэтилоксаминоат и некоторые другие производные адамантана типа 1-AdCH 2 OCH 2 CH(OH)CH 2 NRR΄ обладают анестезирующим действием .
Антибактериальное действие, сравнимое с антибактериальным препаратом 5-нитро-8-гидроксихинолином, оказывают N-(нитрофенил)-адамантил-карбоксамиды и адамантилзамещённые N-(1-метилпиридиний)йодиды .
Перфторированный адамантан применяется в качестве компонента искусственной крови . Есть данные об антиагрегационной способности производных адамантана по отношению к различным путям агрегации тромбоцитов .
1.4. Производные адамантана, влияющие на агрегацию тромбоцитов
Ярко выраженная липофильная природа каркасного углеродного фрагмента адамантана обуславливает высокую биологическую активность его производных. Введение в молекулу производного высоколипофильного радикала усиливает его взаимодействие с гидрофобными областями рецепторных молекул .
Показано влияние модифицированных адамантильных и амидных фрагментов амидов N-[(адамантоил-1)-фенил]-антраниловой кислоты на 5-НТ 2 рецепторы тромбоцитов человека: снижение их АДФ- и серотонин-индуцированной агрегации . Кроме того, известно, что сродство к данным рецепторам обусловлено наличием объёмного адамантильного заместителя . Была предположена биологическая активность ряда 1-адамантанкарбоксамидов в качестве антагонистов 5-НТ 2 рецепторов тромбоцитов. Высокоспецифическое сродство к изучаемым рецепторам и мощный антиагрегационный эффект был подтверждён и invitro и invivo для (S)-N-{1-пирролидин-3-ил}-1-адамантанкарбоксамид гидрохлорид гидрата (10-(S), Y-39241) .
Ряд синтезированных адамантильных производных гетерофункциональных соединений проявил свойства антагонистов и синергистов серотонина при исследовании их влияния на серотонин-индуцированную агрегацию тромбоцитов человека .
Глава 2. Материал и методы исследования
Проведено исследование влияния на индуцированную агрегацию тромбоцитов человека 19-ти производных адамантана с различающимися по структуре и свойствам заместителями (табл. 1).
Таблица 1.
Исследуемые соединения
| № | Химическая формула вещества |
Название вещества |
| 1 | N-(1-адамантил)-3-аминопиридина гидрохлорид | |
| 2 | 2--1-метилпиридиний иодид | |
| 3 | 4--1-метилпиридиний иодид | |
| 4 | N-(1-метилпиридиний-3-ил)-адамантил-1-карбоксамид иодид | |
| 5 | 2--4-бром-1-метилпиридиний иодид | |
| 6 | 3‑(адамантил-1-карбонилокси)-2‑этил‑6-метилпиридин | |
| 7 | 2‑(адамантил‑1‑карбонилокси)-пиридин |
Продолжение таблицы 1.
| № | Химическая формула вещества |
Название вещества |
| 8 | N-(1-адамантилацетил)-2-аминопиридин | |
| 9 | N-(1-адамантилацетил)-4-аминопиридин | |
| 10 | N-(1-адамантилацетил)-3-аминопиридин | |
| 11 | N-(1-адамантоил)-D,L-триптофан |
|
| 12 | 3-(1-адамантанкарбоксамидо-этил)индол | |
| 13 | N-(1-адамантоил)-D,L-гистидин | |
| 14 | 4-адамантанкарбоксамидо-этилбензоат | |
| 15 | 4-адамантанкарбоксамидо-N- -бензоат | |
| 16 | 4-адамантанкарбоксамидо-N--бензамид |
Продолжение таблицы 1.
Эти вещества синтезированы в лаборатории кафедры органической химии СамГУ научным сотрудником кандидатом химических наук Ермохиным В. А. . Строение вновь синтезированных соединений доказано данными ЯМР 1 Н и ИК спектроскопии (Приложение, табл. 1-4). ИК спектры соединений снимали на спектрофотометре ИКС-29 в таблетках из KBr. Спектры ЯМР получены на приборе BrukerWP-200 SY (рабочая частота 200,13 МГц). В качестве растворителя применяли ДМСО-d 6 , CD 3 CN, CDCl 3 . Отсчёт химических сдвигов проводили относительно сигнала ТМС. Для подтверждения индивидуальности всех полученных соединений проводили тонкослойную хроматографию на пластинках SilufolUV в этилацетате. Соединения № 18 и № 19 были синтезированы ранее .
Агрегация тромбоцитов исследована фотометрическим методом Борна . Принцип метода основан на регистрации снижения оптической плотности исследуемой плазмы после введения в неё тромбоцитагрегирующего агента. Происходит «склеивание» тромбоцитов, образующиеся агрегаты постепенно оседают и проба плазмы становится более прозрачной.
Биологический материал предоставлен самарской областной станцией переливания крови. Кровь, взятую из локтевой вены, стабилизировали 3,8 % раствором цитрата натрия (9:1). Богатую тромбоцитами плазму получали центрифугированием цитратной крови при 1000 об/мин в течение 10 минут. Плазму разбавляли 0,154 М раствором хлорида натрия, рН 7,2 до достижения оптической плотности пробы 0,4-0,6 ед. (стандартизация плазмы). Плазму разливали в кюветы по 2 мл. Изучаемые соединения растворяли в 96 % этиловом спирте. В качестве агрегирующих агентов использовали растворённый в 0,154 М физ. растворе рН 7,2 5-гидрокситриптамин креатин сульфат (серотонин) и АДФ в конечной концентрации 1*10 -5 М. В эксперименте в качестве агреганта использовался также адреналина гидрохлорид в концентрации 0,1 %. Конечные концентрации всех соединений составляли 0,099 мг/мл. Оптическую плотность измеряли против контроля на СФ-46 при длине волны 600 нм, кювета 10 мм (в контрольную пробу добавляли спирт вместо раствора вещества) в течение 30 минут. Об интенсивности агрегации судили по изменению оптической плотности относительно контроля, которое выражали в условных единицах: ∆ А 30 (изменение оптической плотности за 30 минут)´1000=х у. е.
Полученные данные анализировали для установления значений их достоверности согласно критерию Стьюдента. Данные считали достоверными при уровне значимости р<0,05 .
Глава 3. Результаты и обсуждение
Биологическая активность любого химического соединения определяется его структурой, конформацией его молекулы. В данном эксперименте сначала были выявлены некоторые производные адамантана, влияющие на серотонин-индуцируемую агрегацию тромбоцитов человека invitro. Затем были исследованы механизмы влияния этих соединений на агрегацию тромбоцитов человека.
3.1. Влияние производных адамантана на серотонин-индуцируемую агрегацию тромбоцитов человека in vitro
Адамантилсодержащие производные гидрокси- и аминопиридинов и иодидов N -адамантоил-1-метилпиридиния и О‑адамантоил- N -метилпиридиния. В пробах с добавлением производного адамантана № 1 агрегация тромбоцитов менее интенсивна по сравнению с агрегацией в контрольных пробах, но это отличие не достоверно (табл. 2).
Таблица 2.
Влияние N-(1-адамантил)-3-аминопиридина гидрохлорида на серотонин-индуцированную агрегацию тромбоцитов человека invitro
Соединение № 2 отличается от соединения № 1 положением азота аминопиридина и наличием метильной группы у этого атома азота, заряд азота пиридина в соединении № 2 экранирован йодид-ионом, а в соединении № 1 – хлорид-ионом (табл. 1). Несмотря на такие незначительные структурные различия, соединение № 2 проявило себя как синергист серотонина в процессе агрегации тромбоцитов (табл. 3).
Таблица 3.
Влияние 2--1-метилпиридиний иодида на серотонин-индуцированную агрегацию тромбоцитов человека invitro
<0,001 к контролю.
Из полученных данных был сделан вывод о том, что способность изучаемых соединений влиять на серотонин-индуцируемую агрегацию тромбоцитов коррелирует с положением азота пиридина по отношению к кислороду адамантоильной группировки. Вероятно, азот пиридина во втором положении придаёт производному адамантана свойства синергиста серотонина. Для проверки этого предположения был поставлен эксперимент с веществом, азот пиридина которого стоит в четвёртом положении (табл. 4).
Таблица 4.
Влияние 4--1-метилпиридиний иодида на серотонин-индуцированную агрегацию тромбоцитов человека invitro
Предположение подтвердилось: в пробах с 4--1-метилпиридиний иодидом интенсивность агрегации оказалась почти такой же, как и в контрольных пробах. Производное № 3 не влияет на серотонин-индуцируемую агрегацию тромбоцитов, хотя отличается от производного № 2 только положением метилированного азота. Чтобы проверить, зависит ли способность соединения № 2 влиять на серотонин-индуцируемую агрегацию тромбоцитов от наличия в адамантоильной группировке аминогруппы, поставили эксперимент с веществом с карбоксильной группой (табл. 5).
Таблица 5.
Влияние N-(1-метилпиридиний-3-ил)-адамантил-1-карбоксамид иодида на серотонин-индуцированную агрегацию тромбоцитов человека invitro
П р и м е ч а н и е: уровень достоверности результатов ***- p<0,001 к контролю.
Результаты эксперимента с веществом № 4 почти такие же, как и с веществом № 2, что ещё раз доказывает зависимость биологической активности производных адамантана именно от положения азота пиридина по отношению к кислороду адамантоильной группировки.
Модификация адамантоильной группировки метилированием и внедрение в молекулу производного бромид-иона существенно снизило способность полученного соединения влиять на серотонин-индуцируемую агрегацию тромбоцитов человека (табл. 6).
Таблица 6.
Влияние 2--4-бром-1-метилпиридиний иодида на серотонин-индуцированную агрегацию тромбоцитов человека invitro
Также “неудачной” оказалась модификация соединения, в результате которой получилось более гидрофобное производное № 6. 3‑(адамантил-1-карбонилокси)-2‑этил‑6-метилпиридин (соединение № 6), в отличие от предыдущих веществ, не имеет внутримолекулярного заряда, и почти не проявил биологической активности в отношении агрегации тромбоцитов (табл. 7).
Таблица 7.
Влияние 3‑(адамантил-1-карбонилокси)-2‑этил‑6-метилпиридина на серотонин-индуцированную агрегацию тромбоцитов человека invitro
Молекула 2‑(адамантил‑1‑карбонилокси)-пиридина тоже сильно гидрофобна, и тоже почти не повлияла на индуцируемую серотонином агрегацию тромбоцитов (табл. 8).
Таблица 8.
Влияние 2‑(адамантил‑1‑карбонилокси)-пиридина на серотонин-индуцированную агрегацию тромбоцитов человека invitro
И N-(1-адамантилацетил)-2-аминопиридин почти не влияет на серотонин-индуцируемую агрегацию тромбоцитов, хотя азот пиридина находится во втором положении (табл. 9).
Таблица 9.
Влияние N-(1-адамантилацетил)-2-аминопиридина на серотонин-индуцированную агрегацию тромбоцитов человека invitro
Значит, если азот пиридина в составе молекулы производного адамантана находится во втором положении, для проявления его биологической активности необходима полярность молекулы, способствующая её гидратации.
Интересно, что N-(1-адамантилацетил)-4-аминопиридин оказался синергистом серотонина (табл. 10).
Таблица 10.
Влияние N-(1-адамантилацетил)-4-аминопиридинана серотонин-индуцированную агрегацию тромбоцитов человека invitro
П р и м е ч а н и е: уровень достоверности результатов ٭- p<0,05 к контролю.
Подобное аминопиридиновое производное адамантана с азотом пиридина в третьем положении не влияет на серотониновую агрегацию тромбоцитов (табл. 11).
Таблица 11.
Влияние N-(1-адамантилацетил)-3-аминопиридина на серотонин-индуцированную агрегацию тромбоцитов человека invitro
Из всех гидрофобных адамантил-аминопиридинов синергистическими серотонину свойствами обладает соединение, азот пиридина которого находится в четвёртом положении (табл. 10). Из более гидрофильных адамантильных (адамантоильных) пиридиновых производных синергистами серотонина оказались аминопиридиновые производные, азот пиридина которого находится во втором положении (табл. 3, 5). Заметим, что эти соединения имеют внутримолекулярный заряд, который облегчает их растворимость в водной межклеточной среде, сохраняя при этом мембранотропность.
N -адамантоил-α-аминокислоты. Рацемическая смесь триптофанового производного адамантана проявила слабую тенденцию к замедлению серотонин-индуцируемой агрегации тромбоцитов (табл. 12).
Таблица 12.
Влияние N-(1-адамантоил)-D,L-триптофанана серотонин-индуцированную агрегацию тромбоцитов человека invitro
Триптаминовое производное способно ускорять серотонин-индуцируемую агрегацию тромбоцитов, а смесь из D- и L-гистидиновых производных напротив, достоверно её замедляет (табл. 13, 14).
Таблица 13.
Влияние 3-(1-адамантанкарбоксамидо-этил)индолана серотонин-индуцированную агрегацию тромбоцитов человека invitro
П р и м е ч а н и е: уровень достоверности результатов ٭٭٭- p<0,001 к контролю.
Таблица 14.
Влияние N-(1-адамантоил)-D,L-гистидинана серотонин-индуцированную агрегацию тромбоцитов человека invitro
П р и м е ч а н и е: уровень достоверности результатов ٭٭٭- p<0,001 к контролю.
Из экспериментальных данных видно, что смеси D- и L-аминокислотных производных адамантана проявляют антагонистические серотонину свойства в процессе агрегации тромбоцитов, а триптаминовое производное – синергистические. Способность триптаминового производного (3-(1-адамантанкарбоксамидо-этил)индола) усиливать индуцируемую серотонином агрегацию тромбоцитов, скорее всего, связана с его структурным сходством с самим серотонином (рис.7).
Рис. 7. Строение серотонина (а) и 3-(1-адамантанкарбоксамидо-этил)индола (б). Видно сходство структуры серотонина (5-гидрокситриптамина) и биологически активного производного адамантана.
Адамантилсодержащие производные 4-аминобензойной кислоты. Первым из производных 4-аминобензойной кислоты был исследован 4-адамантанкарбоксамидо-этилбензоат, который проявил тенденцию к снижению интенсивности серотонин-индуцируемой агрегации тромбоцитов (табл. 15).
Таблица 15.
Влияние 4-адамантанкарбоксамидо-этилбензоата на серотонин-индуцированную агрегацию тромбоцитов человека invitro
Модификация 4-адамантанкарбоксамидо-этилбензоата, в результате которой получился 4-адамантанкарбоксамидо-N- -бензоат, привела к появлению у нового соединения синергистических серотонину свойств (табл. 16).
Таблица 16.
- -бензоата на серотонин-индуцированную агрегацию тромбоцитов человека invitro
П р и м е ч а н и е: уровень достоверности результатов ٭٭٭- p<0,001 к контролю.
Модификация бензоата до бензамида приводит к изменению биологической активности производного вплоть до тенденции к замедлению индуцируемой серотонином агрегации тромбоцитов в опытных пробах по сравнению с контролем (табл. 17).
Таблица 17.
Влияние 4-адамантанкарбоксамидо-N- -бензамида на серотонин-индуцированную агрегацию тромбоцитов человека invitro
При хлорировании и оксиметилировании бензамида получается производное адамантана, обладающее свойствами синергиста серотонина (табл. 18).
Таблица 18.
Влияние 4-адамантанкарбоксамидо-5-хлор-N--2-метоксибензамида на серотонин-индуцированную агрегацию тромбоцитов человека invitro
П р и м е ч а н и е: уровень достоверности результатов ٭- р<0,05 к контролю.
Адамантилсодержащие производные этилендиамина. Ещё два производных адамантана, которые теоретически должны влиять на серотонин-индуцируемую агрегацию тромбоцитов, в эксперименте не проявили ожидаемой биологической активности. В опытных пробах с добавлением N-(1-адамантоил)этилендиамина гидрохлорида почти такая же интенсивность агрегации тромбоцитов как и в контрольных пробах (табл. 19).
Таблица 19.
Влияние N-(1-адамантоил)этилендиамина гидрохлоридана серотонин-индуцированную агрегацию тромбоцитов человека invitro
Производное адамантана № 19, молекула которого симметрична и включает две адамантоильных группировки, проявило лишь тенденцию к снижению интенсивности индуцируемой серотонином агрегации тромбоцитов (табл. 20).
Таблица 20.
Влияние N,N"-ди-(1-адамантоил)этилендиаминана серотонин-индуцированную агрегацию тромбоцитов человека invitro
Для серотонина на мембране тромбоцитов существует два типа рецепторов – ионные каналы и рецепторы, сопряжённые с G-белками . Поэтому серотонин способен индуцировать процессы агрегации тромбоцитов двумя путями, являясь лигандом для двух разных типов рецепторов. На функционирование какого типа серотониновых рецепторов влияют добавляемые в опытные пробы соединения, можно проверить аналогичными экспериментами, где агрегантом является известный лиганд для рецепторов, сопряжённых с G-белками, например, АДФ и адреналин. Для этих экспериментов было отобрано 5 соединений: соединения № 2, 12, 9, проявившие наиболее выраженное синергистическое серотонину действие (соединения № 15 и 17 больше не исследовались, так как они оказались неустойчивыми); соединение № 13, проявившее свойство антагониста серотонина и № 14, которое хоть очень слабо, но тоже замедляет серотонин-индуцируемую агрегацию тромбоцитов.
Активность вещества № 4 почти не отличалось от таковой вещества № 2, их структура тоже мало отличается (табл. 3, 5), поэтому чтоб не усложнять эксперимент, вещество № 4 не исследовалось.
Более чётко влияние изучаемых соединений на серотонин-индуцируемую агрегацию тромбоцитов видно на графиках зависимости оптической плотности проб от времени.
Снижение оптической плотности в опытных пробах с добавлением вещества № 2 происходит интенсивнее, чем в контрольных, что говорит о способности этого соединения усиливать агрегацию тромбоцитов. В опытных пробах с добавлением вещества № 9 оптическая плотность тоже снижается интенсивнее, чем в контрольных, хотя здесь отличие опыта от контроля более сглажено. Вещество № 12 тоже усиливает серотонин-индуцируемую агрегацию тромбоцитов. Влияние на агрегацию тромбоцитов вещества № 13 замедляет снижение оптической плотности опытных проб. Вещество № 14 тоже способно замедлять серотонин-индуцируемую агрегацию тромбоцитов, но менее интенсивно, чем вещество № 13.
3.2. Механизмы влияния производных адамантана на индуцируемую агрегацию тромбоцитов человека in vitro
К настоящему моменту в литературе отсутствует информация о возможных механизмах влияния подобных соединений на функционирование тромбоцитарных рецепторов. Влияние веществ на скорость агрегации тромбоцитов может быть обусловлено их способностью усиливать сродство серотонина к рецепторам (синергисты) или каким-либо образом блокировать доступ лиганда к своему рецептору. Также возможным является агонистическое влияние на рецептор с малой константой ассоциации агониста с рецепторной молекулой. Для проверки данных предположений были поставлены эксперименты с веществами без индуцирования агрегации известными агрегантами. Исследования показали, что вещества не способны самостоятельно индуцировать агрегацию (табл. 21, рис 10).
Таблица 21.
Влияние производных адамантана на не индуцированную агрегацию тромбоцитов человека invitro
Рис. 10. Динамика снижения оптической плотности проб при не индуцируемой агрегации тромбоцитов человека: (а б в г д ) В опытные пробы добавляли соединение № 14. Интенсивность агрегации в опытных пробах почти не отличается от таковой в контрольных пробах.
Полученные данные свидетельствует о том, что исследуемые производные не способны самостоятельно проводить сигнал, а влияют лишь на сродство серотонина к рецептору тромбоцита, модулируя эффективность проведения сигнала активации (рис. 8).
Из результатов экспериментов с добавлением веществ при индукции агрегации тромбоцитов АДФ, представленных в таблице 22, видно, что 4-адамантанкарбоксамидо-этилбензоат (соединение № 14) способен значительно замедлять АДФ-индуцированную агрегацию тромбоцитов. Возможно, эта его способность обусловливает незначительное замедление и серотонин-индуцированной агрегации.
Таблица 22.
Влияние производных адамантана на АДФ-индуцированную агрегацию тромбоцитов человека invitro
<0,05; ٭٭- p<0,01 к контролю.
Очевидно, производное № 14 влияет на рецепторы тромбоцитов, сопряжённые с G-белками, блокируя проведение сигнала активации агрегации. А N-(1-адамантилацетил)-4-аминопиридин (производное № 9), напротив, интенсивно ускоряет проведение сигнала и усиливает АДФ-индуцированную агрегацию тромбоцитов. 2--1-метилпиридиний иодид (производное № 2) здесь тоже усиливает агрегацию, но не так интенсивно. Вещество № 13 не влияет на АДФ-индуцированную агрегацию тромбоцитов. Результат действия индольного производного (№ 12) несколько размыт высокой ошибкой среднего значения из-за индивидуальных колебаний показателей плазмы разных людей. Но ускорение агрегации тромбоцитов в пробах с веществом № 12 по сравнению с контролем достоверно (табл. 22).
Результаты экспериментов с добавлением веществ при индукции агрегации тромбоцитов АДФ показаны на рисунке 11.
Рис. 11. Динамика снижения оптической плотности проб при АДФ-индуцируемой агрегации тромбоцитов человека. (а ) В опытные пробы добавляли соединение № 2. (б ) В опытные пробы добавляли соединение № 9. (в ) В опытные пробы добавляли соединение № 12. (г ) В опытные пробы добавляли соединение № 13. (д ) В опытные пробы добавляли соединение № 14.
Заметим, что влияние исследуемых соединений на АДФ-индуцируемую агрегацию тромбоцитов наблюдается не сразу после их добавления в пробы (как в эксперименте с серотонином), а лишь после определённого латентного периода (рис. 11). Это значит, что соединения начинают влиять на рецепторы для АДФ только после внедрения в плазматическую мембрану, изменяя их сродство к лиганду.
Снижение оптической плотности в опытных пробах с соединением № 2 в течение первых 20 минут мало отличается от такового в контрольных, но за последующие 10 минут оно более интенсивно и достоверно отличается от контроля. После пяти минут экспозиции падение оптической плотности в пробах с добавлением вещества № 9 интенсивнее. Отличия динамики снижения оптической плотности опытных и контрольных проб в эксперименте с веществом № 12 также начинаются после пяти минут экспозиции. Интенсивность снижения оптической плотности в опытных пробах в эксперименте с веществом № 13 почти не отличается от таковой в контрольных. Соединение № 14 способствует замедлению падения оптической плотности опытных проб после 10 минут от начала агрегации (рис. 11).
Полученные данные говорят о том, что, скорее всего, производне № 2 и 13 изменяют сродство серотонина к рецепторам-ионным каналам, а производные № 9, 12 и 14 способны влиять на сродство лиганда к своим гликопротеиновым рецепторам (табл. 22).
К гликопротеиновым рецепторам, функционирующим посредством G-белков, относится большинство из множества всех рецепторов клеток (в данном случае тромбоцитов). Возникает вопрос о специфичности влияния исследуемых производных адамантана. Возможно, среди данных соединений присутствуют синергисты других агрегантов, также проводящих сигнал активации посредством ассоциации с гликопротеиновыми рецепторами. Действительно, предположение подтверждено экспериментом с добавлением веществ при индукции агрегации тромбоцитов адреналином: вещества № 9 и № 12 ускоряют проведение сигнала и через сопряжённые с G-белком адренорецепторы, а вещества № 2, 13 и 14 не способны модулировать функционирование тромбоцитарных адренорецепторов (табл. 23, рис. 12).
Таблица 23.
Влияние производных адамантана на адреналин-индуцированную агрегацию тромбоцитов человека invitro
П р и м е ч а н и е: уровни достоверности результатов ٭- р<0,05; ٭٭٭- p<0,001 к контролю.
Динамика снижения оптической плотности в пробах с добавлением соединения № 2 не отличается от таковой в контрольных (так же и в эксперименте с соединениями № 13 и № 14). Значит, эти соединения не способны влиять на проведение сигнала активации через адренорецепторы. Добавление в опытные пробы соединения № 9 значительно ускоряло падение их оптической плотности по сравнению с контролем, то же наблюдалось и при добавлении соединения № 12.
Совершенно очевидно, что под влиянием соединений № 9 и 12 увеличивается эффективность того этапа (или нескольких этапов) функционирования гликопротеиновых рецепторов, который является общим для всех типов этих рецепторов, не зависимо от их специфичности (рис. 12, б, в).
Таким образом, доказанная предыдущими исследованиями биологическая активность аминопиридиновых производных адамантана подтверждается и в нашем эксперименте: N-(1-адамантилацетил)-4-аминопиридин (№ 9) значительно увеличивает сродство тромбоцитарных рецепторов ко всем используемым в эксперименте агрегантам (табл. 10, 22, 23). Причём это, скорее всего, свидетельствует о его способности к модуляции функционирования гликопротеиновых рецепторов. Модификация аминопиридинового заместителя в составе производного адамантана приводит к изменению его активности: 2--1-метилпиридиний иодид (№ 2) усиливает, видимо, сродство серотонина к своим рецепторам со свойствами ионных каналов, почти не влияя на сродство к гликопротеиновым рецепторам (табл. 3, 22, 23).
Исследованиями показана способность объёмных заместителей, вносимых в молекулу адамантана, изменять сродство серотонина к своим рецепторам. Существуют также экспериментально подтверждённые данные о том, что введение в молекулу производного высоколипофильного радикала усиливает его взаимодействие с гидрофобными областями рецепторных молекул . Эти данные подтверждаются и в нашем эксперименте: 4-адамантанкарбоксамидо-этилбензоат (№ 14) действительно имеет подобную активность (табл. 15, 22), способствуя замедлению проведения сигнала активации АДФ и серотонином. Причём некоторое блокирование проведения сигнала под действием 4-адамантанкарбоксамидо-этилбензоата наблюдается через 10 минут. На адреналин-индуцируемую агрегацию он почти не влияет, и причину этого явления ещё предстоит выяснить.
Аминокислотные производные адамантана – 3-(1-адамантанкарбоксамидо-этил)индол (№ 12) и N-(1-адамантоил)-D,L-гистидин (табл. 1, № 13), – тоже имеют интересную активность. N-(1-адамантоил)-D,L-гистидин, вероятно, антагонистически действует только на серотониновые ионные каналы тромбоцитов, а 3-(1-адамантанкарбоксамидо-этил)индол весьма эффективно ускоряет агрегацию тромбоцитов, индуцированную и серотонином, и АДФ, и адреналином (табл. 13, 22, 23), т. е. его активность направлена на рецепторы гликопротеинового типа. Вещество № 12 интересно ещё и тем, что его структура очень похожа на структуру самого серотонина (рис. 7). Несмотря на то, что вещество № 12, как и серотонин, является производным триптофана, оно не является его агонистом (табл. 21).
Заключение
Влияние производных адамантана на способность тромбоцитов человека к агрегации – одно из многих проявлений их биологической активности. Высокая липофильность и мембранотропность адамантана, а так же возможность внесения в его структуру разнообразных заместителей, позволяют синтезировать соединения с заданной биологической активностью. Но, как и все высоко активные вещества, производные адамантана токсичны, так как даже соединения с хорошо изученным биохимическим механизмом действия способны вызывать побочные реакции организма. Поскольку в фармакологическую практику всё чаще стали внедряться лекарственные препараты на основе производных адамантана, детальное изучение их биохимических свойств становится необходимым. Результаты исследований, приведённые в данной работе, возможно, являются первыми шагами на пути к созданию высокоэффективных лекарств, позволяющих решить проблемы многих людей, страдающих патологиями системы гемостаза.
Скорее всего, биологическая активность производных адамантана по отношению к серотониновым рецепторам – ионным каналам – обусловлена способностью изменять их конформацию за счёт изменения структуры плазматической мембраны. Тем более это вероятно, если принять во внимание мембранотропность адамантанового ядра и канальное строение серотониновых рецепторов. Поскольку другие серотониновые рецепторы как и рецепторы для АДФ и для адреналина являются гликопротеинами, и их строение более сложно и менее зависит от ориентации фосфолипидных слоёв мембраны, здесь более вероятен механизм модуляции функционирования самого рецептора, в результате чего активация G-белков происходит эффективнее.
Проведённые исследования позволили сделать новые выводы относительно зависимости влияния производных адамантана на агрегацию тромбоцитов человека от химической структуры адамантильных заместителей. Прояснились некоторые вопросы, связанные с механизмами воздействия вновь синтезированных соединений на рецепторы тромбоцитов человека: очевидна их способность изменять сродство лигандов не только к рецепторам – ионным каналам, но и к рецепторам, сопряжённым с G-белком, что определяет направление дальнейших исследований.
Выводы
1.Влияние производных адамантана на серотонин-индуцируемую агрегацию тромбоцитов человека определяется их структурой и взаимным расположением атомов в молекуле.
2.Производные адамантана оказывают различное действие на индуцируемую агрегацию тромбоцитов человека, что является следствием их способности влиять на функционирование разных тромбоцитарных рецепторов, но ни одно из исследуемых соединений не способно самостоятельно индуцировать агрегацию тромбоцитов (не имеют свойств агонистов).
3.Усиливающее влияние 2--1-метилпиридиний иодида наиболее выражено при серотонин-индуцируемой агрегации тромбоцитов, что обусловлено его способностью усиливать сродство серотонина к своим рецепторам со свойствами ионных каналов, почти не влияя на сродство к гликопротеиновым рецепторам (синергист серотонина).
4.N-(1-адамантилацетил)-4-аминопиридин и 3-(1-адамантанкарбоксамидо-этил)индол усиливают агрегацию тромбоцитов, индуцируемую и серотонином, и адреналином, и АДФ, увеличивая эффективность одного из этапов функционирования гликопротеиновых рецепторов, общего для всех типов этих рецепторов.
5.N-(1-адамантоил)-D,L-гистидин, напротив, эффективно замедляет серотонин-индуцируемую агрегацию тромбоцитов, блокируя проведение сигнала активации серотонином (антагонист серотонина).
6.4-адамантанкарбоксамидо-этилбензоат способствует замедлению агрегации тромбоцитов, индуцируемой и АДФ, и серотонином, не влияя при этом на адреналин-индуцируемую агрегацию.
Литература
1.Андронова В. Л. Противовирусная активность препаратов ряда адамантана. //Антибиотики и химиотерапия. - 1996. - 41, № 718. - С. 26-30.
2.Антоников И. М. Форма тромбоцитов крысы на различных этапах процесса их агрегации. // Вестн. Акад. мед. наук. -1981. - 55, № 6. С. 78-79.
3.Багрий Е. И. Адамантаны: получение, свойства, применение. - М.: Наука, 1989. - 264 с.
4.Балуда В. П., Баркаган З. С., Гольдберг Е. Д. и др. Лабораторные методы исследования системы гемостаза. - Томск, 1980. - 84 с.
5.Балуда В. П., Лукоянова Т. И. Простациклин-генерирующая система стенки сосудов и тромбогенез. // 1 Всесоюзная конференция, Поражение сосудистой стенки и гемостаз. (Полтава, 1981). - С. 20.
6.Белушкина Н. Н., Северина И. С. Торможение АДФ-индуцируемой агрегации тромбоцитов гуанидинотиолами – новым классом активаторов гуанилатциклазы и субстратов NO-синтазы. // Биохимия. - 1996. - 61, № 12. - С. 2140-2146.
7.Вотяков В. И., Бореко Е. И., Владыко Г. В., Карако Н. И., Коробченко Л. В. Поиск соединений, активных в отношении вирусов гриппа и возбудителей других ОРВИ. // Химиотерапия и химиопрофилактика гриппа и ОРЗ. (Сб. науч. трудов). - Л., 1990. - С.150.
8.Воробьёв В. Б. Физиология гемостаза. - Ростов-на-Дону: Изд. дом Профф. Пресс, 2004. - 192 с.
9.Врынчану Н. А. Острая токсичность новых производных адамантана и норборнана. // Токсикология лекарств. – Киев: Институт фармакологии и токсикологии АМН Украины, 2001. – С. 20-25.
10. Голубева М. Г. Роль адренорецепторов в регуляции системы свёртывания крови. // Физиология человека. - 1989. - 15, № 6. С. 127-137.
11. Даниленко Г. И., Мохорт М. А., Триус Ф. П. Синтез и биологическая активность производных адамантана. N-(адамантоил-1)антраниловые кислоты. // Хим.-фарм. Журн. - 1973. - 17, №10. С. 15-17.
12. Даниленко Г. I., Рибалко С. Л., Максимов Ю. М. та інш. Пошук інгібіторів ВІЛ та герпесу серед похідних каркасних сполук та піридинкарбонових кислот // Биополимеры и клетка. - 1999. - 15, № 3 – С. 207-212.
13. Ермохин В. А. Синтез и биологическая активность адаманитлпроизводных гетерофункциональных ароматических аминов и азотсодержащих гетероциклов: Автореф. дис. ... канд. химич. наук. – Самара, 2007. - 22 с.
14. Ермохин В. А., Зарубин Ю. П., Пурыгин П. П., Золотарёв П. Н. Синтез и антибактериальная активность N-(нитрофенил)-адамантил-1-карбоксамидов и адамантансодержаших иодидов N-метилпиридиния. // Вестник СамГУ. Естественнонаучная серия. – 2007. – 56, № 6. – С. 378-384.
15. Ермохин В. А., Пурыгин П. П., Зарубин Ю. П. Адамантановые производные эфиров и амидов 4-аминобензойной кислоты. // Вестник СамГУ. Естественнонаучная серия. – 2006. – 49, № 9. – С. 92-96.
16. Ермохин В. А., Пурыгин П. П., Клёнова Н. А. Синтез и гемолитическая активность N-адамантоилзамещённых гетероциклических аминов и анилинов. // Вестник СамГУ. Естественнонаучная серия. – 2004. – 34, № 4. – С. 138-144.
17. Зинченко В. П., Долгачева Л. П. Внутриклеточная сигнализация. – Пущино: Электронное издательство «Аналитическая микроскопия», 2003. – 84 с.
18. Индулен М. К., Колнина В. А., Рязанцева Г. М., Бубович В. И. Механизмы антивирусного действия производных адамантана. Под ред. М. К. Индулен. - Рига: Знание, 1981. – 168 с.
19. Исаев С. Д., Новикова М. И., Коробенко Л. В. Синтез и биологическая активность адамантилсодержащих оксазолов и имидозолов. // Физиологически активные вещества: Республиканский межведомственный сборник. – 1991. – 125, № 23. – С. 26-31.
20. Касьян Л. И., Касьян А. О., Голодаева Е. А. // Журнал органической химии. – 2000. – 36, № 12. - С. 1776-1779.
21. Ковалёв И. Е., Щипулина Н. В. // Доклады Российской академии наук. - 2001. - 378, № 6. - С. 819-822.
22. Козинец Г. И., Сарычева Т. Г. и др. Атлас клеток крови и костного мозга. Под ред. Г. И. Козинца. – М.: «Триада-Х», 1998. - 160 с.
23. Колосова Е. А., Курбатова С. В., Соловова Н. В., Финкельштейн Е. Е., Яшкин С. Н. Топология полиэдрических молекул на примере производных адамантана. // Вестник СамГУ. Естественнонаучная серия. – 2001. – 20, № 2. – С. 178-188.
24. Красуцкий П. А., Семёнов И. П., Новикова М. Н., Весёлова Т. В. Аминокислоты ряда адамантана. Синтез и противовирусная активность α-аминокислот ряда адамантана и их производных. // Хим-фарм. Журнал. - 1985. - 57, № 7. - С. 17-25.
25. Кубарь О. И. Анализ побочных реакций, вызываемых противовирусными препаратами // Безопасность лекарств. – 1997. – 78, № 1. - С. 5-7.
26. Кульберг А. Я. Рецепторы клеточных мембран. Биохимия мембран. Под ред. А. А. Болдырева. Кн. 4. – М.: Высш. шк., 1987. – 103 с.
27. Курбатова С. В., Шумская Н. Ю. Исследование взаимосвязи между структурой и хроматографическим поведением карбонильных производных адамантана в условиях обращённо-фазовой ВЭЖХ. // Вестник СамГУ. Естественнонаучная серия. - 2004. –34, № 6. – С. 158-168.
28. Литвинов В. П. Биологическая активность производных адамантана. // Химия гетероциклических соединений. - 2002. - 50, № 1. - С. 12-39.
29. Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека: в 2-х томах. Т. 2. Пер. с англ.: - М.: Мир, 1993. – 415 с.
30. Морозов И. С., Петров В. И., Сергеева С. А. Фармакология адамантанов. - Волгоград: Волгоградская мед. академия, 2001. - 320 с.
31. Ноздрачев А. Д., Баженов Ю. И., Баранникова И. А., Батуев А. С. и др. Начала физиологии: Учебник для ВУЗов. Под ред. Акад. А. Д. Ноздрачева. - СПб.: «Лань», 2001. – 1088 с.
32. Панченко Е. П., Добровольский А. Б. Тромбозы в кардиологии. Механизмы развития и возможности терапии. – М.: Спорт и культура, 1999. – 464 с.
33. Пархоменко А. Н. Патофизиология острого тромбоза в венечных артериях сердца: представления о патогенезе острого коронарного синдрома. – Киев: Инст. кардиологии им. Н. Д. Стражеско АМН Украины, 2004. – 25 с.
34. Покровский В. М., Коротько Г. Ф., Наточин Ю. В. и др. Физиология человека. Учебник в 2-х томах. Под ред. В. М. Покровского, Г. Ф. Коротько. – М.: Медицина, 1997. – 368 с.
35. Русяев В. А., Вотяков В. И., Федоров А. Н. Сравнительная оценка фармакотерапевтического эффекта соединения Д-84 и ремантадина при экспериментальной гриппозной инфекции белых мышей // Антивирусная активность и механизм действия различных химических соединений (под ред. Р. А. Кукайн). - Рига: Зинатне, 1979. - С. 49-53.
36. Самаль А. Б., Черенкевич С. И., Хмара Н. Ф. Агрегация тромбоцитов: методы изучения и механизмы. – М.: Университетское, 1990. - 104 с.
37. Севастьянова В. В., Краюшкин М. М., Юрченко А. Г. Успехи химии адамантана. // Успехи химии. – 1970. – 39, № 10. – С. 1721-1754.
38. Сергеев П. В., Сейфулла Р. Д, Майский А. И Физико-химические механизмы и гормональная регуляция свёртывания крови. – М: Высш. шк., 1974. - 145 с.
39. Столяров З. Е., Лященко К. П. Аминопроизводные адамантана и иммунный ответ // Хим-фарм. журнал. – 1995. – 8, № 3. – С. 9-10.
40. Трифонов Е. В. Психофизиология человека. – М.: Высш. шк., 2004. – 130 с.
41. Фролов Ю. П. Математические методы в биологии. ЭВМ и Программирование. - Самара: «Самарский университет», 1997. - 265 с.
42. Циммерман М., Ениг В., Вутке В. и др. Физиология человека. В 3-х томах. Т. 2. Пер. с англ. Под ред. Р. Шмидта и Г. Тевса. – М.: Мир, 1996. – 313 с.
43. Чирков Ю. Ю., Белушкина Н. Н., Тыщук И. А., Северина И. С. Изменения в активности гуанилатциклазы тромбоцитов человека при АДФ-индуцируемой агрегации. // Бюллетень экспериментальной биол. и медицины. – 1991. – 52, № 2. – С. 152-154.
44. Bojarski A. J., Mokrosz M. J. et al. The influence of substitution at aromatic part of 1,2,3,4-tetrahydroisoquinoline on in vitro and in vivo 5-HT1A/5-HT2A receptor activities of its 1-adamantoyloaminoalkyl derivatives. // J. Bioorganic and Med. Chem. – 2002. – 10, N 5. Р. 87-95.
45. Bonaventure P., Nepomuceno D., Miller K., Chen J., Kuei C., Kamme F., and oth. Molecular and pharmacological characterization of seretonin 5-HT 2A and 5- HT 2B receptor subtypes in dog. // Europen Journal of Pharmacology. - 2005. - 513, N 3. Р. 181-192.
46. Cerrito E., Lazzaro M. P., Gaudio E., Arminio P. and Aloisi G. 5HT 2 -receptors and serotonin release: Their role in human platelet aggregation. // Life Sciences. - 1993. -53, N 3. Р. 209-215.
47. Clerck F., Xhonneux B., Wiele r. Biochemical mechanisms in 5-hydroxytriptamine-induced human platelet aggregation // Agents Actions.- 1985. - 17, N 2. Р. 220-228.
48. Cohen M. L., Robertson D. W., Bloomquist W. E. and Wilson H. C. LY215840, a potent 5-hydroxytryptamine (5-HT) 2 receptor antagonist, blocks vascular and platelet 5-HT 2 receptors and delays occlusion in a rabbit model of thrombosis. // J. Pharmacol. Exp. Ther. - 1992.- 261, N 5. Р. 202-208.
49. Crawford N., Castle A. Microtudule and contractile submit proteins of blood platelets. Role in haemostatic activities. // Microtubules and microtubule inhibitors. – 1975. – 305, N 6. Р. 229-246.
50. Dahl M., Puusten T., Uotila P. The effect of arachidonic acid on the aggregability of human platelet rich plasma. // Prostaglandins leicotriens Med. – 1982. – 8, N 2. Р. 133-139.
51. F. F. De Clerk and A. G. Herman. 5-hydroxyryptamine and platelet aggregation. // Fed. Proc. – 1983. –42, N 3. Р. 228-232.
52. Fred De Clerck, Benoit Xhonneux, Josée Leysen and Paul A. J. Janssen. The involvement of 5-HT 2 -receptor sites in the activation of cat platelets. // Thrombosis Research. – 1984. – 33, N 1. Р.305-321.
53. Frojmovic M. M., Milton J. C. Human platelet size, shape and related functions in health and diasease. // Physol. Rev. – 1982. – 62, N 2. Р. 185-201.
54. Frojmovic M. M., Milton J. C. Physical. Chemical and functional changes following platelet activation in normal and “giant” platelet. // Blood Cells. - 1983. - 9, N 5. Р. 359-382.
55. Fujio M., Kuroita T. et. al. N--1-adamantanecarboxamides as Novel 5-HT2 Receptor Antagonists. // Bioorganic and Med. Chem. Lett. – 2000. – 4, N 10. Р. 2457-2461.
56. Fujio M., Togo Y., Tomozane H., Kuroita T., Morio Y., Katayama J. and Matsumoto Y. N-{methyl}cyclohexane-carboxamides as selective 5-HT 1A receptor agonists. // Bioorganic and Medicinal Chemistry Lett. – 2000. - 10, N 6. Р. 509-512.
57. Gear A., R. L. Preaggregation reactions of platelets. // Blood. – 1981. – 58, N 3. Р. 477-490.
58. Hoyer D., Clarke D.E. and Fozard J.R. et al. International Union of Pharmacology classification of receptors for 5-hydroxytryptamine (Serotonin). // Pharmacol Rev. – 1994. – 10, N 46.Р. 157.
59. Jacobs K. H. Determination of the turn-off reaction for the epinefrine-ingibited human platelet adenylate cyclase. // Eur. J. Biochem. – 1983. – 132, № 1. Р. 125-130.
60. Komiotis D., Pananookooln S. J., Zaw K., Dieter J. P., G. C. Le Breton and D. L. Venton. Synthesis of dl-cis- and (4R,5R)-trans-7--5(Z)-heptenoic acid analogues as platelet thromboxane A 2 receptor antagonist. // European Journal of Medicinal Chemistry. – 1995. – 30, N 4. Р. 321-326.
61. Kroll M. H., Schafer A. I. Biochemical mechanisms of platelet activation. // Blood. – 1989. – 74, N 9. Р. 1181-1195.
62. Li Hui Yang, Demetra Callas, Jean Marc Herbert and Jawed Fareed. Modulation of platelet function and vascular smooth muscle contractile actions by a novel, selective, highly potent 5-HT 2
antagonist (SR46349). //
Thrombosis Research. - 81, N 5. P. 555-567.
63. Li N., Wallen N.H., Ladjevardi M. and Hjemdahl P. Effects of serotonin on platelet activation in whole blood. // Blood Coagul. Fibrinolysis. – 1997. -8, N 27. Р. 517-523.
64. Mahaut-Smith M. P.; Ennion S. J.; Rolf M. G.; Evans R. J. ADP is not an agonist at P2X(1) receptors: evidence for separate receptors stimulated by ATP and ADP on human platelets. // Br. J. Pharmacol. – 2000. – 131, N 1. Р. 10-14.
65. Marlar R. A., Kleiss A. J., Griffin J. H. An alternative extrinsic pathway of human blood coagulation. // Blood. – 1982. – 15, N 60. Р. 13-53.
66. Maurer-Spurej E., Pittendreigh C. and Solomons K. The influence of selective serotonin reuptake inhibitors on human platelet serotonin. // Thromb. Haemost. – 2004. – 12, N 91. Р. 119-128.
67. Nagatomo T., Rashid M., Abul Muntasir H. and Komiyama T. Functions of 5-HT 2A receptor and its antagonists in the cardiovascular system. // Pharmacology and Therapeutics. – 2004. – 104, N 1. Р. 59-81.
68. Niewiarowski S. Platelet release reaction and secreted platelet proteins. // Haemostasis and trombosis. – 1981. – 11, N 18. Р. 73-83.
69. Ogawa T., Sugidachi A., Tanaka N., Fujimoto K. and Asai F. Pharmacological profiles of R-96544, the active form of a novel 5-HT 2A receptor antagonist R-102444. // European Journal of Pharmacology . – 2002. – 457, N 20. P. 107-114.
70. P. Anne McBride, J. John Mann, Margaret J. Polley, Amy J. Wiley and John A. Sweeney. Assessment of binding indices and physiological responsiveness of the 5-HT 2
receptor on human platelets. //
Life Sciences. - 1987. - 40, N 4. Р. 1799-1809.
71. Pauwels P. J. 5-HT receptors and their ligands. // Tocris. Rev. – 2003. – 27, N 25. Р. 1-7.
72. Perri T., Pasini F. L., Frigerio C. et al. Pharmacodynamics of ticlopidine in man in relation to plasma and blood cell concentration. // Europ. J. Clin. Pharmacology. – 1991. – 41, N 20. Р. 429-434.
73. Rao G. H. R., White J. C. Epinephrine potentiation of arachidonate-induce aggregation of cyclooxygenase-deficient platelets. // J. Biol. Chem. – 1980. – 255, N 8. Р. 355-366.
74. Rivail L. et al. Large-scale molecular dynamics of a G protein-coupled receptor, the human 5-HT4 serotonin receptor, in a lipid bilayer. // Journal of Molecular Structure, Theochem. – 2007. – 31, N 48. P. 1-8.
75. Rudnick G., Fishkes H., Nelson P. J., Schuldiner S. Evidence for two dittinct serotonin transport systems in platelets. // J. Biol. Chem. – 1980. – 17, N 2. Р. 3638-3641.
76. Rupprecht H.-J. Adenosine diphosphate receptor antagonists: from pharmacology to clinical practice. // Europ. Heart J. – 2000. – 2, N 7. Р. 1-5.
77. Savi P., Herbert J.-M. ADP receptors on platelets and ADP-selective antiaggregating agents. // Medical Research reviews. – 1996. – 16, N 2. Р. 159-179.
78. Savi P., Heilmann E., Nurden P. et al. Clopidogrel: an antithrombotic drug acting on the ADP-dependent activation pathway of human platelets. // Clin. Appl. Thrombosis. Hemostasis. – 1996. – 2, N 3. Р. 35-42.
79. Smith S. K., Limbird L. E. Solibilization of human platelet α-adrenogetic receptors: evidence that agonist occupancy of the receptor stabilizes receptor-effector interactions. // Proc. Nat. Acad. Sci. USA. 1981. – 78, N7. Р. 4026-4030.
80. Stanford S. C. 5-Hydroxytryptamine. // Neurotransmitters, Drugs and Brain Function. – 2001. – 57, N 45. Р. 187-209.
81. Thebault J. J., Kieffer G., Cariou R. Single-dose pharmacodynamics of clopidogrel. // Semin Thromb Hemost. – 1999. – 25, N 2. Р. 3-8.
82. Thomas D. R. et al. SB-699551-A (3-cyclopentyl-N--N-[(40-{[(2-phenylethyl)amino]methyl}-4-biphenylyl)methyl]propanamide dihydrochloride), a novel 5-ht5A receptor-selective antagonist, enhances 5-HT neuronal function: Evidence for an autoreceptor role for the 5-ht5A receptor in guinea pig brain. // Neuropharmacology. – 2006. – 51, N 5. Р. 566-577.
83. Vickers S. P., Easton N., Malcolm C. S., Allen N. H., Porter R. H., Bickerdike M. J. and Kennett G. A. Modulation of 5-HT 2A receptor-mediated head-twitch behaviour in the rat by 5-HT 2C receptor agonists. // Pharmacology Biochemistry and Behavior. – 2001. – 69, N 8. Р. 643-652.
84. Walcourt-Ambakederemo A. and Winlow W. 5-HT receptors on identified Lymnaea neurones in culture: Pharmacological characterization of 5-HT 2 receptors. // General Pharmacology: The Vascular System. – 1994. – 25, N 6. Р. 1079-1092.
85. White J. G. The secretory pathway of bovine platelets. // Blood. - 1987. – 69, N 3. Р. 878-885.
86. White J. G., Clawson C. C., Gerrarg J. M. Platelet ultrastuctura. // Homeostasis and thrombosis. Edinburg e. a. – 1981. – 65, N 28. Р. 22-49.
87. Mössner R. and Lesch K. P. Role of serotonin in the immune system and in neuroimmune interactions. // Brain Behavior Immun. – 1998. – 75, N 12.Р. 249.
ПРИЛОЖЕНИЕ
Таблица 1.
Данные ИК и ЯМР 1 H спектров для иодидов N-адамантоил-1-метилпиридиния и О‑адамантоил-N-метилпиридиния
|
соединения |
ИК спектр, n, см -1 | J , Гц | |||
| CH 2 | СO–NHR | Остальные | H Ad , H CH 2 | H Ar(Het) , H NH | |
| 1 | 2846, 2912 |
1223, 1176, 1072 |
1.66 c (12H, CH 2), 4.25 c (3H, CH 3) |
8.06 т (1H 5 , Het), J = 4 8.49 д (1H 6 , Het), J =2,7 8.67 д (1H 4 , Het), J = 2 9.43 с (1H 2 , Het), |
|
| 2 | 3300 | 8.00к (2H, Het) 8.42с (1H, Het) |
|||
| 3 | 1.67 c (12H, CH 2), 2.19 c (2H, CH 2), |
8.04 т (1H 5 , Het), J =4 8.41 д (1H 6 , Het), J =2,2 8.67 д (1H 4 , Het), J =1,4 9.38 с (1H 2 , Het), 10.89 c (1H, NH) |
|||
1620,1590, 1520,1395, |
1.67 с (6H, CH 2), 1.96 с (3Н, СН), 3.61 с (3Н, СН 3) |
7.16т (1H 5
, Het), 8.36 м (2H 3 ,4 , Het) |
|||
| 1755 | 1.14 м (3H, CH 3), 1.24 т (2H, CH 2), 1.67 с (3H, CH 3), 1.74 м (12H, CH 2), 2.02 м (3H, CH), |
8.14д (1H 5 , Het), 8.34д (1H 4 , Het) |
|||
Таблица 2.
Данные ИК и ЯМР 1 Н спектров для адамантилсодержащих производныхгидрокси- и аминопиридинов
|
соединения |
ИК спектр, n, см -1 | Спектр ЯМР 1 Н, d (м.д., от ТМС); J , Гц | |||
| CH 2 | СO-NHR | H Ad, H CH2 | H Ar(Het) , H NH | ||
| 3310 | 1630 | 1.70 c (6H, CH 2), 1.92 c (6H, CH 2), |
7.95д (1H, Het), 8.05 д (1H, Het), 8.42 с (1H, Het), |
||
| 1740 ν(С=О), 1590 ν(C=C) Ar, 1215 ν(C–O–C) | 1.67 c (6H, CH 2), 1.80 c (6H, CH 2), 1.96 c (3H, CH) |
6.18 т (1H 5 , Het), 6.31д (1H 6 , Het), 7.36м (2H 3,4 , Het) |
|||
| 8 | 2846, 2900 | 3255 | 1685 | 1.63 c (12H,CH 2), 1.92 c (3H, CH 2), 2.15 c (2H, CH 2) |
7.06 т (1H 4 , Het, J = 3, 7.74 т (1H 5 , Het), J =4, 8.09 д (1H 6 , Het), J =4, 8.28 д (1H 3
, Het), J
=4, |
| 2850, 2900 | 3440, 3230 |
1676 | 1.62 c (12H, CH 2), 1.92 c (3H, CH 2), 2.09 c (2H, CH 2) |
7.57 с (1H 2,6 , Het), 8.38 с (1H 3,5 , Het), 10.09 c (1H, NH) |
|
| 10 | 3370 | 1.63 c (12H, CH 2), 1.94 c (3H, CH 2), 2.10 c (2H, CH 2) |
7.42 д (1H 6 , Het), J =1,4, 8.07 т (1H 5 , Het), J =4, 8.24 д (1H 4 , Het), J =5, 8.78 с (1H 2 , Het), 10.12 c (1H, NH) |
||
Таблица 3.
Данные ИК и ЯМР 1 Н спектров для N-адамантоил-α-аминокислот
|
соединения |
ИК спектр, n, см -1 | Спектр ЯМР 1 Н, d (м.д., от ТМС); J , Гц | ||
| (C–H) Ad | n(N–H), n(C=O) | H Ad, H CH2 | H Ar(Het) , H NH | |
| 2908, 2850 | 3394, 3317, 1720, 1693, 1654 | 1.65м (6H, CH 2), 1.76м (6H, CH 2), 2.80с (1H, СН), 4.15м (2H, CH 2) |
6.96 м (1H, Het), 7.04 м (1H, Het), 7.10 с (1H, Het), 7.27 м (1H, Het), 7.45 м (1H, Het), 7.53 м (1H, NH), 8.40 c (1H, NH, Het) |
|
| 2908, 2850 | 3413, 3217, 1693, 1640 | 1.65 м (6H, CH 2), 1.76 м (6H, CH 2), 1.94 c (3H, CH), 2.79 т (7,78 Гц, 2H, СН 2) 3.30 к (14,6 Гц 2H, СН 2) |
6.96 т (7,0 Гц, 1H, Het), 7.04 т (7,0 Гц, 1H, Het), 7.10 д (4 Гц, 1H, Het), 7.33 д (8 Гц, 1H, Het), 7.41 т (6 Гц, 1H, Het), 7.53 д (8 Гц, 1H, NH), 8.88 c (1H, NH Het) |
|
| 2904, 2847 | 3413, 1700, 1640 | 1.64 м (6H, CH 2), 1.76 д (6H, CH 2), 1.92 c (3H, CH), 2.92 м (2H, CH 2), 3.99 к (11,24 8 Гц, 1H, CH) |
6.59 с (1H, Неt), 6.96 д (6 Гц, 1H, Het), 7.41 д (4 Гц, 1H, NH), 8.43 c (1H, NH Нet) |
|
Таблица 4.
Данные ИК и ЯМР 1 Н спектров для адамантилсодержащих производных 4-аминобензойной кислоты
| № соединения | ИК спектр, n, см -1 | Спектр ЯМР 1 Н, d (м.д., от ТМС); J , Гц | |
| n(N-H), n(C=O), прочие | ν(C-H)Ad | ||
| 3310, 1740, 1680, 1635, 1550, 1280, 850 | 2912, 2800 | 1.70 c (6H, CH 2); 1.91 c (6H, CH 2); 2.01 c (3H, CH); 1.30 т (3H, CH 3), J = 7 Гц; 4.27 д (2H, CH 2), J = 8 Гц; 7.84 д (2H, Ar), J = 8 Гц; 7.87 д (2H, Ar), J = 8 Гц; |
|
| 3410, 3198, 1728, 1670, 1450, 1404, 1272,1126 | 2904, 2851 | 1.30 м (6H, CH 3), 1.70–2.01 м (15H, Ad), 2.48 c (4H, CH 2), 3.27 c (2H, CH 2), 7.84 д (2H, Ar) J=8 Гц, 7.87 д (2H, Ar) J = 8 Гц, 9.36 с (1H, NH) |
|
| 339–-3213 уш, 1693, 1605, 1542, 1508, 1450, 1411, 1284 | 2912, 2851 | 1.27 м (6H, CH 3), 1.76–2.01 м (15H, Ad), 2.47 м (4H, CH 2), 3.47 м (2H, CH 2), 7.21 м (2H, Ar), 7.65 м (2H, Аr), 7.84 c (1H, NH), 9.4 с (1H, NH) |
|
| 3350–3317 уш, 1693, 1605, 1450, 1411, 1284, 1253 | 2912, 2854 | 1.24 c (6H, CH 3), 1.70–2.08 м (15H, Ad), 2.50 м (4H, CH 2), 3.3 м (2H, CH 2), 3.80 c (3H, CH 3), 7,16 c (H, Ar), 7.25 c (H, Ar), 7.59 д (1H, NH), J=2 Гц, 9.68 с (1H, NH) |
|
Камфора относится к производным бициклотептана. Природную -камфору получают из камфорного дерева (Китай, Япония) перегонкой с паром. Рацемическую камфору (3) синтезируют из а-пинена (1) через формиат (2) Она возбуждает центральную нервную систему (ЦНС), стимулирует дыхание и обменные процессы в миокарде (кардиотоник) Назначают ее при сердечной недостаточности, отравлениях наркотиками и снотворными и для растираний при ревматизме Введение атома
брома в -положение к кетонной группе резко меняет фармакологическую картину у производного камфоры. Бромкамфора (4), улучшая сердечную деятельность, приобретает седативные свойства и успокаивает ЦНС. Применяют ее при неврастении и неврозах сердца:

Производные полициклической адамантановой системы предложены в качестве антивирусных средств. 1-Амино-адамантан (8) (мидантан, амантадин) получают бромированием адамантана (5) в присутствии меди до 1-бромадамантана (6), который действием формамида переводят в 1-формил-аминопроизводное (7). Гидролиз последнего в присутствии HCI приводит к мидантану (первое синтетическое антигриппозное средство). Алкилированием аминоадамантана 1-хлорглюк-уроновой кислотой в присутствии основания получают его глюкуронид (9) (глудантан - лекарственное вещество для лечения паркинсонизма и вирусных заболеваний глаз - конъюнктивитов):

(Еще одно антифиппозное лекарственное вещество - ремантадин (13) - синтезируют замещением брома в соединении (6) на карбоксильную группу, действуя муравьиной кислотой в олеуме (эта система генерирует СО , необходимые для заместительного гидроксикарбонилирования). Далее с помощью тионилхлорида превращают кислоту (10) в ее хлорангидрид, который
обрабатывают этоксимагниймалоновым диэфиром и превращают в ацилпроизводное (11). Его без выделения гидролизуют до дикислоты, а последнюю декарбоксилируют и получают 4-ацетиладамантан (12). Соединение (12) затем подвергают восстановительному аминированию в системе формамид/муравьиная кислота, что приводит к ремантадину (13):

Уникальная структура лиганд-связывающих участков семикратно пересекающих мембрану рецепторов позволяет связывать лиганды различной природы и молекулярной массы в широком диапазоне от 32 для Ca2+ до более чем 102 кДа для гликопротеинов.
Большинство обычных низкомолекулярных гормонов (типа адреналина и ацетилхолина) связывается с участками внутри гидрофобного ядра (a). Пептидные и белковые лиганды присоединяются к внешней поверхности рецептора (b, c). Некоторые лиганды низкого молекулярного веса, Ca2+ и аминокислоты (глутамат, ГАМК) связываются с длинными участками на N-конце, индуцируя их переход в новую конформацию, в которой длинный участок взаимодействует с рецептором (d). В случае рецепторов, активируемых отрезающей протеазой (e), новый N-конец действует как автолиганд. Отрезанный пептид может также взаимодействовать с другим рецептором.
1.3. Биологическая активность производных адамантана
Производные адамантана как физиологически активные вещества находят широкое применение с 70-х годов XX века . Сам адамантан (трициклодекан, С10Н16) принадлежит к числу трициклических нафтенов мостикового типа (рис. 6).
Рис. 6. Структура молекулы адамантана.
Молекула его состоит из трёх конденсированных циклогексановых колец в конформации кресла. Пространственная модель молекулы адамантана – высокосимметричная конструкция с малой поверхностью и незначительными силами межмолекулярного взаимодействия в кристаллической решётке . Из всех трициклических углеводородов адамантан наиболее устойчив, что объясняется тетраэдрической направленностью связей всех атомов углерода и их фиксированным положением.
Биологическая активность производных адамантана обусловлена симметрией и объёмностью пространственного строения, значительной липофильностью жёсткого углеводородного каркаса адамантана, что позволяет им легко проникать через биологические мембраны . Поэтому модификация органических соединений с помощью адамантильного радикала значительно изменяет их биологическую активность, нередко усиливая её. С помощью метода спиновых меток было показано, что адамантан, попадая в липидный бислой, способен разрушать гексагональную упаковку метиленовых группировок, характерную для двойного слоя фосфолипидов, и нарушать осевое расположение алкильных цепей фосфолипидов, модифицируя тем самым функциональные свойства клеточных мембран . Принимая во внимание важность порядка расположения метиленовых групп липидов биологических мембран как фактора функционирования связанных с мембранами ферментов, можно отметить косвенное влияние адамантана на их активность.
На данный момент синтезировано более 1000 новых производных адамантана. Фармакологическое изучение показало наличие среди них веществ, обладающих выраженной психотропной, иммунотропной, противовирусной, курареподобной, антикаталептической, противоаллергической активностями, а также соединений, влияющих на ферментативную систему печени . Амиды адамантанкарбоновых кислот проявляют антибактериальную активность .
Есть данные о результатах модификации молекулы энкефалина аминокислотами ряда адамантана. Введённый в положение 5 молекулы энкефалина (S)-адамантилаланин придаёт опиоидному пептиду резистентность по отношению к ферментам, легко разрушающим немодифицированный энкефалин (химотрипсин, проназа, нейтральная протеаза, термолизин) .
Показано, что физиологической активностью обладают азотсодержащие производные. Первым в медицинскую практику в 1966 г. вошёл гидрохлорид 1-аминоадамантана, обладающий противовирусной активностью в отношении штаммов вирусов типа А2, его фирменные названия: мидантан, симметрел, амантадин. Эти препараты используются для профилактики респираторных заболеваний, т. к. обладают способностью блокировать проникновение вируса в клетку. Предполагают, что данные препараты способны работать на начальных этапах репродукции вируса, блокируя синтез вирусоспецифичных РНК . Противовирусная активность некоторых аминопроизводных адамантана связана с их способностью ингибировать ПК С . Ремантадин (полирем, флумадин), как липофильное слабое основание способен повышать рН эндосомального содержимого и препятствовать депротеинизации вируса .
В клинической практике для лечения вирусных заболеваний применяются также такие препараты, как ацикловир (виролекс, герпесин, зовиракс, лизавир, суправиран), диданозин, фоскарнет (триаптен), ганцикловир (цимевен), ламивудин, рибавирин (виразол, рибамидил), ставудин, трифлуридин, видарабин, зальцитабин (хивид), зидовудин (азидотимидин, ретровир) . Однако, у большинства этих препаратов относительно узкий спектр противовирусного действия, их недостатком является наличие разнообразных побочных реакций, появление резистентных штаммов вирусов и др. .
Противовирусной активностью в отношении штаммов вирусов типа А2 обладают и производные алкиладамантанов: 1-гидрокси-3,5диметил-7-этиладамантан, 1-метокси-3,5диметиладамантан, которые, в отличие от мидантана, показали высокую противовирусную активность и в отношении штаммов strains of rhino viruses и herpes simplex . Ряд гидрокси-, галоген- и меркаптопроизводных амидов адамантана тоже обладают противовирусной активностью .
Показано, что амантадин способен препятствовать развитию очагов саркомы на зародышевой культуре, другие производные адамантана могут служить снотворными средствами, антималярийными препаратами, инсектицидами . В экспериментах с использованием ВИЧ-инфицированных человеческих лимфобластоидных клеток показано, что некоторые производные адамантана обладают анти-ВИЧ-активностью . Мидантан используется в неврологической клинике для лечения болезни Паркинсона и паркинсонического синдрома. Подобную активность проявляют хлорангидриды 3,5,7-алкилзамещённых 1-аминоадамантанов, некоторые из которых обладают антагонистическими дофамину свойствами. Некоторые четвертичные аммониевые основания с 2-адамантильным радикалом способны действовать как миорелаксанты периферического действия (курареподобная активность). Производные 1-аминоадамантана и 3,3-диамино-1,1-диадамантила противокаталептически активны, бактериостатическое влияние оказывают адамантанкарбоновые кислоты и фосфаты адамантантиолов и их производных. Диалкиламиновые эфиры адамантантиокарбоновой кислоты проявляют бактерицидную, фунгицидную, гербицидную активность . Натриевая соль β-(1-адамантан)-пропионовой кислоты обладает желчегонным действием. 1-адамантиламмоний-β-хлорэтилоксаминоат и некоторые другие производные адамантана типа 1-AdCH2OCH2CH(OH)CH2NRR΄ обладают анестезирующим действием .
Антибактериальное действие, сравнимое с антибактериальным препаратом 5-нитро-8-гидроксихинолином, оказывают N-(нитрофенил)-адамантил-карбоксамиды и адамантилзамещённые N-(1-метилпиридиний)йодиды .
Перфторированный адамантан применяется в качестве компонента искусственной крови . Есть данные об антиагрегационной способности производных адамантана по отношению к различным путям агрегации тромбоцитов .
 Всеволод большое гнездо как правитель
Всеволод большое гнездо как правитель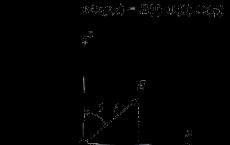 Как заполняются электронные уровни, подуровни и орбитали по мере усложнения атома
Как заполняются электронные уровни, подуровни и орбитали по мере усложнения атома Самые первые спички в мире
Самые первые спички в мире